Метаболический препарат на основе таурина с показаниями к применению:
- хроническая НК I-IIIф.к.;
- интоксикация сердечными гликозидами;
- сахарный диабет.
Влияние содержания таурина в плазме крови и генетической предрасположенности к диабету на изменение чувствительности к инсулину на фоне гипокалорийной диеты (реферат)
Резюме Цель исследования: оценить взаимосвязь между содержанием таурина в плазме крови и снижением резистентности к инсулину при изменении питания у участников исследования POUNDS Lost (Preventing Overweight Using Novel Dietary Strategies - предотвращение избыточного веса с помощью новых диетических стратегий), проанализировать модифицирующее влияние таурина на генетическую предрасположенность к сахарному диабету 2 типа (СД 2). Материал и методы: 811 человек с лишним весом или ожирением рандомизированы в зависимости от диеты с различным содержанием жиров, белков и углеводов: группа 1 - 20%, 15% и 65%; группа 2 - 20%, 25% и 55%; группа 3 - 40%, 15% и 45%; группа 4 - 40%, 25% и 35% соответственно. Масса тела, обхват талии и уровень глюкозы натощак измеряли утром перед завтраком исходно, через 6 мес. и через 2 года. Резистентность к инсулину оценивали с помощью индекса HOMA-IR, секрецию инсулина - с помощью индекса HOMA-B. У 711 человек проанализированы генетические данные и результаты измерения уровня таурина в плазме крови. Результаты исследования: у испытуемых с наименьшим терцилем индекса генетического риска СД 2 исходное содержание таурина ассоциировалось с менее выраженным снижением уровня глюкозы натощак и HOMA-IR (р=0,02), а у испытуемых с наибольшим терцилем исходное содержание таурина ассоциировалось с более выраженным снижением уровня инсулина и HOMA-IR (р=0,04). Среди испытуемых с наименьшим и средним терцилем исходного содержания таурина взаимосвязь между генетической предрасположенностью к СД и уменьшением резистентности к инсулину статистически достоверно изменялась на протяжении 2 лет (р=0,05). Среди испытуемых с наибольшим терцилем эта ассоциация статистически достоверно не изменялась на протяжении 2 лет (р=0,26). Заключение: в зависимости от существующего генетического фона и исходного содержания циркулирующего таурина гипокалорийная диета может оказывать благоприятное воздействие на инсулинорезистентность различной выраженности. Ключевые слова: сахарный диабет, инсулин, глюкоза, инсулинорезистентность, гипокалорийная диета, таурин, Дибикор. Для цитирования: Влияние содержания таурина в плазме крови и генетической предрасположенности к диабету на изменение чувствительности к инсулину на фоне гипокалорийной диеты (реферат). РМЖ. 2019;1:1-7.
Plasma taurine and diabetes genetic predisposition effects on insulin sensitivity change in response to very-low-calorie diet (report)
Abstract Aim: to assess the relationship between the plasma taurine level and decreased insulin resistance when changing diet in POUNDS Lost (Preventing Overweight Using Novel Dietary Strategies) study subjects. Adding that, to analyze the taurine modifying effect on type 2 diabetes mellitus (DM 2) genetic predisposition. Patients and Methods: 811 overweight or obese subjects were randomized on the basis of 4 diets with different fat, protein, and carbohydrate levels: group 1 - 20%, 15% and 65%; group 2 - 20%, 25% and 55%; group 3 - 40%, 15% and 45%; group 4 - 40%, 25% and 35% respectively. Body weight, waist circumference, and fasting glucose were measured in the morning before breakfast at baseline, after 6 months and after 2 years. Insulin resistance was estimated by homeostasis model assessment of insulin resistance (HOMA-IR), insulin secretion - by homeostasis model assessment of в-cell function (HOMA-B). Genetic data and results of plasma taurine level were analyzed in 711 subjects. Results: in subjects with DM 2 genetic risk score (GRS) across the lowest tertile, the baseline taurine level was associated with a less pronounced decrease in fasting glucose and HOMA-IR (p=0.02). Whereas, a more pronounced decrease in insulin and HOMA-IR (p=0.04) was associated in subjects with baseline taurine level across the highest tertile. Among the subjects with the baseline taurine level across the lowest and medium tertile, the relationship between DM 2 GRS and a decrease in insulin resistance statistically significantly changed over 2 years (p=0.05). Across the highest tertile - statistically significantly did not change over 2 years (p=0.26). Conclusion: depending on the existing genetic background and the baseline taurine level, very-low-calorie diet can have a beneficial effect on insulin resistance of varying severity. Keywords: diabetes mellitus, insulin, glucose, insulin resistance, low-calorie diet, taurine, Dibicor. For citation: Plasma taurine and diabetes genetic predisposition effects on insulin sensitivity change in response to very-low-calorie diet (report). RMJ. 2019;1:1-7.
Введение
Таурин (2-аминоэтансульфоновая кислота) - доминирующая свободная аминокислота, которая составляет примерно 0,1% общей массы тела. Таурин поступает с пищей (мясо, морепродукты), а также может синтезироваться в организме из метионина и цистеина [1]. Метаболизм таурина непосредственно взаимосвязан с развитием ожирения, резистентности к инсулину и диабета. Результаты экспериментальных исследований на животных свидетельствуют о потенциальном положительном влиянии добавок с таурином на уровень глюкозы в крови и резистентность к инсулину [3, 4]. И хотя у больных сахарным диабетом (СД) польза пищевых добавок с таурином все еще подвергается сомнению [5, 6], при СД имеет место пониженное содержание таурина в крови [7-9]. По данным крупного эпидемиологического исследования, уровень выделения таурина с мочой (как мера оценки поступления таурина с пищей) обратно пропорционально взаимосвязан с сердечно-сосудистыми факторами риска [2].
Все больше данных указывает на то, что положительное влияние может быть опосредовано модификацией экспрессии генов ожирения [10-12], метаболизма липидов [13, 14] и диабета [15, 16]. Кроме того, выдвинуто предположение о том, что на риск возникновения заболевания влияют генетическая восприимчивость в сочетании с метаболическим статусом [17]. Авторам не удалось обнаружить исследований, в ходе которых изучалось влияние генетической восприимчивости и уровня циркулирующего таурина на риск развития СД у человека.
Цель исследования: оценить взаимосвязь между содержанием таурина в плазме крови и снижением резистентности к инсулину с позиций изменения характера питания (макронутриентов) у участников крупномасштабного рандомизированного исследования POUNDS Lost (Preventing Overweight Using Novel Dietary Strategies - предотвращение избыточного веса с помощью новых диетических стратегий) продолжительностью 2 года. Анализировалось модифицирующее влияние таурина на генетическую предрасположенность к сахарному диабету 2 типа (СД 2).
Материал и методы
Дизайн исследования
Исследование POUNDS Lost - это рандомизированное интервенционное исследование, в ходе которого сравнивали влияние вариантов гипокалорийной диеты с различным содержанием жиров, белков и углеводов на массу тела. Исследование проводилось в период с 2004 г. по 2007 г. Дизайн исследования, методы и результаты описаны ранее [18]. 811 человек с лишним весом или ожирением (средний возраст 51±9 лет; 64% женщин; ИМТ 33±4 кг/м2) были рандомизированы по 4 группам, в которых была назначена диета с различным содержанием жиров, белков и углеводов: группа 1 - 20%, 15% и 65%; группа 2 - 20%, 25% и 55%; группа 3 - 40%, 15% и 45%; группа 4 - 40%, 25% и 35% соответственно. Все участники исследования POUNDS Lost давали добровольное информированное согласие в письменном виде.
В ходе исследования POUNDS Lost масса тела и обхват талии измеряли утром перед завтраком исходно, через 6 мес. и через 2 года. Уровень глюкозы натощак определяли в аналогичные сроки. Резистентность к инсулину оценивали с помощью индекса HOMA-IR (математическая гомеостатическая модель резистентности к инсулину), который рассчитывали следующим образом:
уровень инсулина натощак (µЕд\мл) х уровень глюкозы натощак(мг/дл)
Содержание таурина натощак определяли в апреле 2014 г. в образцах плазмы крови, хранившихся при температуре -80°С [20, 21].
Генотипирование
ДНК экстрагировали из лейкоцитарной пленки центрифугированной крови с помощью набора QIAmp Blood Kit (Qiagen). Отобран 31 единичный нуклеотидный полиморфизм (SNP), на полногеномном уровне достоверно ассоциированный с развитием СД у лиц европеоидной расы [22]. Для каждого индивидуума авторы суммировали количество аллелей риска SNP, чтобы получить невзвешенный индекс генетического риска [23]. SNP, включенные в индекс генетического риска, были успешно генотипированы у 734 из 811 участников исследования. В доступных образцах ДНК частота успешного генотипирования составила 99%. Реплицированные образцы контроля качества (10%) были включены в каждую плашку генотипирования с конкордантностью более 99% [24]. В настоящее исследование были включены 711 человек, у которых исходно имелись как генетические данные, так и результаты измерения уровня таурина в плазме крови.
Статистический анализ
В качестве первичных исходов оценивали изменения уровней глюкозы натощак, инсулина натощак, инсулинорезистентности и секреции инсулина за период наблюдения. Исходные данные представлены в виде «среднее ± стандартное отклонение». Исходные характеристики сравнивали с использованием теста χ2 в случае категориальных переменных и обобщенных линейных моделей в случае непрерывных переменных. Влияние генетических факторов и изменения характера питания на исходы через 6 мес. и 2 года анализировали с помощью метода генерализованных оценивающих уравнений (GEE). Поправка на ковариаты для ретроспективных исходов в модели 1 осуществлялась на возраст, половую и расовую принадлежность, характер диеты, продолжительность ее соблюдения и исходную величину HOMA-IR. В модели 2 поправка осуществлялась на потерю веса. Потенциальную взаимосвязь между уровнем таурина и индексом генетического риска, а также между уровнем таурина и изменением характера питания оценивали путем включения соответствующих мультипликативных факторов в модели GEE.
В ходе вторичного анализа с помощью смешанных линейных моделей проверяли, меняется ли существенным образом характер влияния генетических факторов на снижение резистентности к инсулину в группах. В ходе анализа чувствительности эти ассоциации оценивали только у представителей европеоидной расы (~ 80% от всех испытуемых); выяснилось, что результаты одинаковы во всей популяции (данные не представлены).
Все представленные величины р были двусторонними. Различия считались статистически достоверными при p<0,05. Статистический анализ проводился с использованием пакета программного обеспечения SAS (версия 9.4).
Результаты исследования
Исходные характеристики испытуемых
Исходные характеристики испытуемых в соответствии с терцилями исходного содержания таурина представлены в таблице 1. Медианы уровня таурина в соответствии с терцилями составили 1,19 µмоль/л, 1,63 µмоль/л и 2,09 µмоль/л. Распределение генетической предрасположенности и характер распределения участников исследования по группам в зависимости от диеты по терцилям уровня таурина статистически достоверно не отличались (р>0,05).
Таблица 1. Исходные характеристики участников исследования POUNDS Lost1 (n = 711) в соответствии с терцилями исходного содержания таурина
Параметр
Терцили исходного уровня таурина
Т1 (n=237)
Т2 (n=237)
Т3 (n=237)
Возраст, лет
50,44±9,11
51,46±9,16
50,86±9,57
Женщины, n (%)
152 (64,1)
132 (55,7)
151 (63,7)
Этническая принадлежность, n (%) евразийская афро-американская латиноамериканская, азиатская и др.
188 (79,3) 36 (15,2) 13 (5,5)
186 (78,5) 39 (16,5) 12 (5,1)
193 (81,4) 33 (13,9) 11 (4,6)
Диета с пониженным содержанием жиров, n (%)
138 (58,2)
110 (46,4)
121 (51,1)
Диета со средним содержанием белков, n (%)
117 (49,4)
124 (52,3)
134 (56,5)
Индекс генетического риска СД 2,n аллелей риска
34,05±4,21
33,86±4,19
34,09±4,75
Исходное содержание таурина, µмоль/л
1,19 (0,3)
1,63 (0,2)
2,09 (0,3)
Рост, см
168,00±8,77
169,12±8,81
168,74±8,83
Исходный ИМТ, кг/м2
32,34±3,63
32,69±4,03
32,92±3,93
Исходный вес, кг
91,65±14,9
93,86±15,77
94,12±15,84
Исходный уровень глюкозы натощак, мг/дл
91,22±12,16
92,78±12,13
91,28±10,69
Исходный уровень инсулина натощак, µЕд/л
9,80 (6,5)
10,60 (9,7)
11,10 (9,1)
Исходная величина HOMA-IR
2,17 (1,7)
2,48 (2,4)
2,51 (2,2)
Исходная величина HOMA-B
125,33 (95,9)
131,20 (98,7)
144,77 (124,9)
1Данные представлены в виде среднее ± стандартное отклонение или n (%), за исключением уровня таурина, инсулина натощак, HOMA-IR и HOMA-B, которые представлены в виде медианы (межквартильный размах). Для категориальных переменных использован критерий χ2, для непрерывных переменных (за исключением исходного уровня таурина) - обобщенная линейная регрессия с ковариатами возраста, половой принадлежности и веса. Статистически достоверных отличий не выявлено (p>0,05).
Влияние содержания таурина в плазме крови и генетической предрасположенности на уменьшение резистентности к инсулину
После внесения поправки на возраст, половую и расовую принадлежность, характер питания, продолжительность соблюдения диеты и исходную величину HOMA-IR оказалось, что взаимосвязь между исходным содержанием таурина и изменением резистентности к инсулину не достигла статистически достоверного уровня (ß=-0,0125; р=0,47).
Количество аллелей риска в наименьшем, среднем и наибольшем терцилях индекса генетического риска (по 31 SNP) оказалось в пределах 15-31, 32-35 и 36-62 соответственно. Установлено, что больший индекс генетического риска ассоциирован с менее выраженным снижением резистентности к инсулину (р=0,03) и уровня глюкозы (р=0,008) при изменении характера питания по сравнению с меньшей генетической предрасположенностью к СД.
Влияние взаимосвязи между исходным содержанием таурина в плазме крови и генетической предрасположенностью на изменение резистентности к инсулину
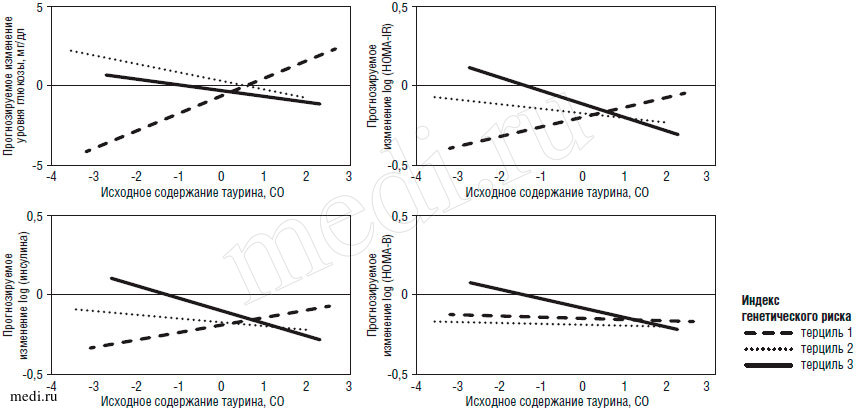
В таблице 2 представлена взаимосвязь между исходным уровнем таурина и изменениями веса, содержания глюкозы натощак, резистентности к инсулину (HOMA-IR) и секреции инсулина (HOMA-B) по терцилям индекса генетического риска СД 2 через 6 месяцев и 2 года. Медианы исходного уровня таурина в соответствии с терцилями индекса генетического риска составили 1,65 µмоль/л, 1,58 µмоль/л и 1,63 µмоль/л соответственно (для линейного тренда р=0,47). В модель 1 внесены поправки на возраст, расовую и половую принадлежность, продолжительность соблюдения диеты и исходную величину для оценки ретроспективного исхода. В модель 2 дополнительно внесена поправка на снижение веса. В обеих моделях ассоциации были одинаковыми, причем даже после внесения поправки на снижение веса сохранялось выраженное влияние ассоциации таурин - индекс генетического риска на изменения уровня глюкозы, инсулина и HOMA-IR. Рисунок 1 иллюстрирует ассоциации между исходным содержанием таурина и прогнозируемыми изменениями гликемических параметров в соответствии с терцилями индекса генетического риска в модели 2. В модели 2 у испытуемых с наименьшим терцилем индекса генетического риска СД 2 исходное содержание таурина ассоциировалось с менее выраженным снижением уровня глюкозы натощак и HOMA-IR (для обоих показателей р=0,02), а у испытуемых с наибольшим терцилем индекса генетического риска СД 2 исходное содержание таурина - с более выраженным снижением уровня инсулина и HOMA-IR (для обоих показателей р=0,04). При этом значения р для ассоциации таурин - индекс генетического риска составили 0,04, 0,01 и 0,002 для изменений уровня глюкозы, инсулина и HOMA-IR соответственно (табл. 2). Статистически достоверного влияния ассоциации таурин - индекс генетического риска на HOMA-B не выявлено.
Таблица 2. Взаимосвязи между исходными уровнями таурина и изменениями массы тела, содержания глюкозы и инсулина, HOMA-IR и HOMA-B в соответствии с терцилями индекса генетического риска СД
Индекс генетического риска диабета
Р
Терциль 1 (наименьший)
Терциль 2
Терциль 3 (наибольший)
Исходное содержание таурина, µмоль/л
1,65(0,7)
1,58 (0,6)
1,63 (0,7)
Характеристики исходов
ß ± СО
Р
ß ± СО
Р
ß ± СО
Р
Модель 1
Изменения массы тела, кг
-0,15±0,44
0,73
-0,47±0,38
0,21
-0,28±0,41
0,49
0,87
Изменения уровня глюкозы, мг/дл
1,00±0,47
0,03
-0,61±0,42
0,15
-0,47±0,57
0,41
0,04
Изменения log инсулина
0,04±0,03
0,14
-0,02±0,03
0,40
-0,06±0,03
0,03
0,006
Изменения log HOMA-IR
0,05±0,03
0,06
-0,03±0,03
0,32
-0,07±0,03
0,03
0,004
Изменения log HOMA-B
-0,02±0,03
0,47
-0,004±0,03
0,89
-0,05±0,03
0,08
0,57
Модель 2
Изменения уровня глюкозы, мг/дл
1,01±0,45
0,02
-0,33±0,43
0,44
-0,35±0,50
0,48
0,04
Изменения log инсулина
0,04±0,02
0,08
0,006±0,02
0,78
-0,05±0,03
0,04
0,01
Изменения log HOMA-IR
0,05±0,02
0,02
0,004±0,02
0,87
-0,06±0,03
0,04
0,002
Изменения log HOMA-B
-0,02±0,03
0,46
0,02±0,03
0,55
-0,04±0,03
0,17
0,68
В первой строке данные представлены в виде медиан (межквартильный размах), в остальных - в виде ß ± стандартное отклонение с поправкой каждого стандартного отклонения на исходное содержание таурина. В модель 1 внесены поправки на возраст, половую и расовую принадлежность, разновидность диеты, продолжительность наблюдения и исходные показатели. В модель 2 также внесена поправка на изменение массы тела. HOMA-IR - математическая гомеостатическая модель резистентности к инсулину, HOMA-B - математическая гомеостатическая модель ß-клеток.
Рис. 1. Взаимосвязь между исходным содержанием таурина и прогнозируемыми изменениями уровней глюкозы и инсулина, инсулинорезистентности и секреции инсулина в соответствии с терцилями генетического риска диабета. В значения р внесена поправка на возраст, половую и расовую принадлежность, разновидность диеты, продолжительность наблюдения, снижение массы тела и исходные показатели (р<0,05)
Изменение резистентности к инсулину
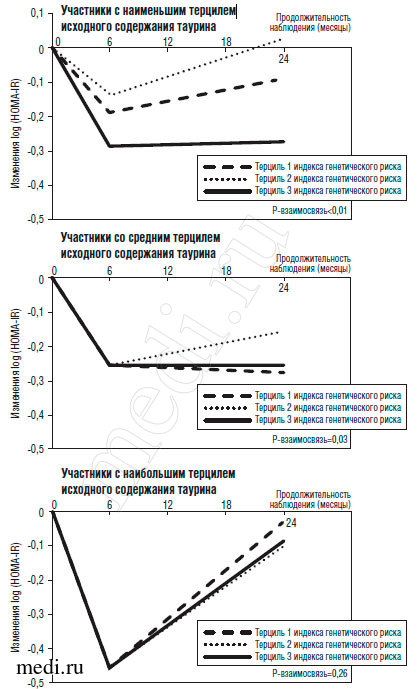
Для оценки траектории изменения резистентности к инсулину за время соблюдения диеты в группах с различным содержанием таурина и различными величинами индекса генетического риска были использованы смешанные линейные модели. Как уже сообщалось ранее [18], наиболее выраженное снижение резистентности к инсулину наблюдается через 6 мес., а через 2 года происходит регресс до исходного уровня. Такая регрессия отчасти может быть связана с несоблюдением диеты и возвратом к прежнему весу. Среди испытуемых с наименьшим и средним терцилем исходного содержания таурина взаимосвязь между генетической предрасположенностью к СД и уменьшением резистентности к инсулину статистически достоверно изменялась на протяжении 2 лет (р=0,05 для взаимосвязи индекс генетического риска - время) (рис. 2). Среди испытуемых с наибольшим терцилем исходного содержания таурина эта ассоциация статистически достоверно не изменялась на протяжении 2 лет (р=0,26 для взаимосвязи индекс генетического риска - время) (рис. 2).
Рис. 2. Изменения инсулинорезистентности по терцилям исходного содержания таурина и генетической предрасположенности к диабету. Внесены поправки на возраст, половую и расовую принадлежность, разновидность диеты, снижение массы тела и исходное значение HOMA-IR. Р-взаимосвязь отражает значение р для влияния взаимосвязи между индексом генетического риска диабета и продолжительностью наблюдения (в месяцах) на изменение HOMA-IR
Обсуждение
В ходе рандомизированного интервенционного исследования по снижению веса путем изменения характера питания продолжительностью 2 года было установлено, что исходное содержание таурина в плазме крови статистически достоверно коррелирует с генетической предрасположенностью к СД 2 с позиций уменьшения резистентности к инсулину. Полученные данные свидетельствуют о том, что среди лиц с повышенным генетическим риском СД исходно более высокое содержание циркулирующего таурина оказывает более благотворное влияние с позиций снижения резистентности к инсулину на фоне диеты, направленной на снижение веса, по сравнению с исходно более низким содержанием таурина. Напротив, в случае меньшего генетического риска СД исходно более низкий уровень таурина оказывает более благотворное влияние, чем его более высокий уровень. При этом взаимосвязь между таурином и генетической предрасположенностью к СД 2 не влияла на секрецию инсулина.
Полученные нами данные о модифицирующем влиянии исходного содержания таурина в крови на генетические факторы, определяющие резистентность к инсулину у человека, являются новыми, однако они не противоречат уже длительно существующей концепции взаимодействия ген - метаболит [17, 25]. Потенциальные механизмы положительного влияния таурина на патогенез СД - модуляция фосфорилирования белка IRS-1 (который ассоциирован с индексом генетического риска) [26] и содержания PPARa (транскрипционного фактора ядерного рецептора, затрагивающего метаболизм жирных кислот), взаимодействие с рецептором инсулина с повышением чувствительности к инсулину [26, 28], влияние на антиоксидантную систему/ воспаление [29] и регуляция экспрессии генов, ассоциированных с развитием СД [9, 15, 16, 30]. Среди этих регулирующих генов отдельного упоминания заслуживает ген рецептора у, активируемого пероксисомными пролифераторами (PPARG) [31], который включен в индекс генетического риска СД 2. Описаны различные механизмы, посредством которых метаболиты осуществляют «тонкую настройку» экспрессии генов, в частности, рибосвитчи, прямое взаимодействие с транскрипционными факторами, регуляция кофакторов, ремоделирование хроматина, модификация хроматина и гормональная сигнализация [32]. Пока не установлено, на каком уровне взаимодействуют клеточные пути, регулируемые генетическими факторами риска развития СД 2 и индуцируемые под действием циркулирующего таурина. Но принимая во внимание представленные выше данные, можно предположить, что снижение резистентности к инсулину под влиянием взаимодействия таурин - индекс генетического риска по крайней мере отчасти отражает регулирующее действие таурина плазмы крови на экспрессию генов, ассоциированных с СД.
Еще одно возможное объяснение заключается в том, что способность таурина препятствовать ожирению [29] может опосредовать взаимодействие ген - таурин с позиций снижения резистентности к инсулину. Впрочем, этой гипотезе противоречит то, что мы наблюдали в ходе нашего исследования, а именно тот факт, что изменение массы тела лишь в незначительной степени затрагивало эффекты взаимодействия ген - таурин. Для уточнения характера подобных функциональных механизмов необходимы новые экспериментальные и клинические исследования.
Для оценки общей вероятности развития СД мы подсчитали генетический риск на основании 31 доказанного варианта, предрасполагающего к СД 2, которые были идентифицированы в ходе полногеномных исследований. Как мы и предполагали, выяснилось, что повышенный генетический риск ассоциирован с менее контролируемым уровнем глюкозы в крови и менее выраженным снижением резистентности к инсулину на фоне гипокалорийной диеты. Среди испытуемых с меньшим содержанием таурина более выраженное уменьшение резистентности к инсулину отмечено у лиц с меньшим генетическим риском, чем у лиц с повышенным генетическим риском (рис. 2). Подобные наблюдения могут быть использованы в целях персонализированной медицины. Впрочем, в повседневной клинической практике приходится обращать внимание на стоимость генетических и/или метаболомических исследований с позиций эффективности подобного индивидуализированного лечения, особенно в группах высокого риска.
Уровень циркулирующего таурина отчасти контролируется за счет изменения скорости его биосинтеза в печени, где он также утилизируется посредством конъюгации с желчными кислотами. Не исключено, что поступающие с пищей жиры способны влиять на содержание циркулирующего таурина путем модуляции экскреции желчных кислот. Так, в ходе опытов на животных установлено, что аминокислоты, поступающие с пищей, могут влиять на метаболизм таурина [33]. Впрочем, мы не отметили какого-либо выраженного влияния диеты (как изменения характера питания в целом, так и конкретного варианта) на уровень циркулирующего таурина (данные не представлены) или влияния взаимосвязи между таурином и составом макронутриентов на инсулинорезистентность. Также не выявлено влияния взаимосвязи между таурином и генетической восприимчивостью к диабету на секрецию инсулина, хотя результаты предшествующих исследований позволяют предположить, что таурин улучшает функционирование островков поджелудочной железы [34, 35].
Насколько нам известно, в ходе этого продолжительного рандомизированного исследования была впервые проанализировано влияние взаимосвязи между содержанием циркулирующего таурина и общей генетической предрасположенностью к диабету на инсулинорезистентность. Мощность данного исследования повышается благодаря анализу исходов во множестве временных точек. Кроме того, модель GEE устойчива к выбору коррелирующих структур и гибка с позиций отсутствия каких-то данных по сравнению с другими моделями [36].
Тем не менее очевидно, что данное исследование обладает недостаточной мощностью в плане выявления незначительных ассоциаций между таурином и инсулинорезистентностью или взаимосвязей между таурином и питанием. Кроме того, полученные результаты не могут быть экстраполированы на популяцию в целом в полной мере, поскольку 80% испытуемых составляли лица европеоидной расы.
Заключение
Таким образом, выявлено влияние взаимосвязи между исходным содержанием таурина в крови и генетической предрасположенностях к диабету на инсулинорезистентность в ответ на смену диеты на 2 года с целью похудения. В зависимости от существующего генетического фона и исходного содержания циркулирующего таурина гипокалорийная диета может оказывать благоприятное воздействие на инсулинорезистентность различной выраженности.
Реферат подготовлен редакцией «РМЖ» по материалам статьи: Zheng Y., Ceglarek U., Huang T. et al. Plasma Taurine, Diabetes Genetic Predisposition, and Changes of Insulin Sensitivity in Responseto Weight-Loss Diets. J Clin Endocrinol Metab. 2016 0ct;101(10):3820-3826. D01:10.1210/jc.2016-1760.
ЛИТЕРАТУРА
Ide T., Kushiro M., Takahashi Y. et al. mRNA expression of enzymes involved in taurine biosynthesis in rat adipose tissues. Metabolism. 2002;51:1191-1197.
Sagara M., Murakami S., Mizushima S. et al. Taurine in 24-h Urine Samples Is Inversely Related to Cardiovascular Risks of Middle Aged Subjects in 50 Populations of the World. Adv Exp Med Biol. 2015;803:623-636.
Anuradha C.V., Balakrishnan S.D. Taurine attenuates hypertension and improves insulin sensitivity in the fructose-fed rat, an animal model of insulin resistance. CanJ Physiol Pharmacol. 1999;77:749-754.
Nakaya Y., Minami A., Harada N. et al. Taurine improves insulin sensitivity in the Otsuka Long-Evans Tokushima Fatty rat, a model of spontaneous type 2 diabetes. Am J Clin Nutr. 2000;71:54-58.
Brons C., Spohr C., Storgaard H. et al. Effect of taurine treatment on insulin secretion and action, and on serum lipid levels in overweight men with a genetic predisposition for type II diabetes mellitus. Eur J Clin Nutr. 2004;58:1239-1247.
Xiao C., Giacca A., Lewis G.F. Oral famine but not N-acetylcysteine ameliorates NEFA-induced impairment in insulin sensitivity and beta cell function in obese and overweight, non-diabetic men. Dia- betologia. 2008;51:139-146.
Franconi F., Bennardini F., Mattana A. et al. Plasma and platelet taurine are reduced in subjects with insulin-dependent diabetes mellitus: effects of famine supplementation. Am J Clin Nutr. 1995;61:1115-1119.
Merheb M., Daher R.T., Nasrallah M. et al. Taurine intestinal absorption and renal excretion test in diabetic patients: a pilot study. Diabetes Care. 2007;30:2652-2654.
Ito T., Schaffer S.W., Azuma J. The potential usefulness of famine on diabetes mellitus and its complications. Amino Acids. 2012;42:1529-1539.
Liaset B., Madsen L., Hao Q. et al. Fish protein hydrolysate elevates plasma bile acids and reduces visceral adipose tissue mass in rats. Biochim Biophys Acta. 2009;1791:254-262.
Lin S., Hirai S., Yamaguchi Y. et al. Taurine improves obesity-induced inflammatory responses and modulates the unbalanced phenotype of adipose tissue macrophages. Mol Nutr Food Res. 2013;57:2155-2165.
Kim K.S., Ji H.I., Chung H. et al. Taurine chloramine modulates the expression of adipo kines through inhibition of the STAT-3 signaling pathway in differentiated human adipocytes. Amino Acids. 2013;45:1415-1422.
Murakami S., Yamagishi I., Asami Y. et al. Hypolipidemic effect of famine in stroke-prone spontaneously hypertensive rats. Pharmacology. 1996;52:303-313.
Yokogoshi H., Mochizuki H., Nanami K. et al. Dietary famine enhances cholesterol degradation and reduces serum and liver cholesterol concentrations in rats fed a high-cholesterol diet. J Nutr. 1999;129:1705-1712.
Carneiro E.M., Latorraca M.Q., Araujo E. et al. Taurine supplementation modulates glucose homeostasis and islet function. Nutr Biochem. 2009;20:503-511.
Sirdah M.M. Protective and therapeutic effectiveness of taurine in diabetes mellitus: a rationale for antioxidant supplementation. Diabetes Metab Syndr. 2015;9:55-64.
Doria A., Wojcik J., Xu R. et al. Interaction between poor glycemic control and 9p21 locus on risk of coronary artery disease in type 2 diabetes. JAMA. 2008;300:2389-2397.
Sacks FM, Bray GA, Carey VJ, et al. Comparison of weight-loss diets with different compositions of fat, protein, and carbohydrates. N Engl J Med. 2009; 360:859-873.
Matthews D.R., Hosker J.P., Rudenski A.S. et al. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. 1985;28:412-419.
Brauer R., Leichtle A., Fiedler G. et al. Preanalytical standardization of amino acid and acylcarnitine metabolite profiling in human blood using tandem mass spectrometry. Metabolomics. 2011;7:344-352.
Ceglarek U., Muller P., Stach B. et al. Validation of the phenylalanine/tyrosine ratio determined by tandem mass spectrometry: sensitive newborn screening for phenylketonuria. Clinical chemistry and laboratory medicine: CCLM / FESCC. 2002;40:693-697.
Morris A.P., Voight B.F., Teslovich T.M. et al. Large- scale association analysis provides insights into the genetic architecture and pathophysiology of type 2 diabetes. Nature genetics. 2012;44:981-990.
Qi Q., Meigs J.B., Rexrode K.M. et al. Diabetes genetic predisposition score and cardiovascular complications among patients with type 2 diabetes. Diabetes care. 2013;36:737-739.
Qi Q., Bray G.A., Smith S.R. et al. Insulin receptor substrate 1 gene variation modifies insulin resistance response to weight-loss diets in a 2-year randomized trial: the Preventing Overweight Using Novel Dietary Strategies (POUNDS LOST) trial. Circulation. 2011;124:563-571.
Roede J.R., Uppal K., Park Y. et al. Transcriptome- metabolome wide association study (TMWAS) of maneb and paraquat neurotoxicity reveals network level interactions in toxicologic mechanism. Toxicology Reports. 2014;1:435-444.
Mature J., Kulakowski E.C. Taurine binding to the purified insulin receptor. Biochem Pharmacol. 1988;37:3755-3760.
Schaffer S.W., Shimada-Takaura K., Jong C.J. et al. Impaired energy metabolism of the taurine-deficient heart. Amino Acids. 2015.
Wu N., Lu Y., He B. et al. Taurine prevents free fatty acid-induced hepatic insulin resistance in association with inhibiting JNK1 activation and improving insulin signaling in vivo. Diabetes Res Clin Pract. 2010;90:288-296.
Imae M., Asano T., Murakami S. Potential role of taurine in the prevention of diabetes and metabolic syndrome. Amino Acids. 2014;46:81-88.
Murakami S. The role of taurine in the pathogenesis of obesity. Mol Nutr Pood Res. 2015;59:1353-1363.
Tsuboyama-Kasaoka N., Shozawa C., Sano K. et al. Taurine (2-aminoethanesulfonic acid) deficiency creates a vicious circle promoting obesity. Endocrinology. 2006;147:3276-3284.
Ladumer A.G. Rheostat control of gene expression by metabolites. Mol Cell. 2006;24:1-11.
Trautwein E.A., Hayes К.С. Amino acid interaction with taurine metabolism in cats. Adv Exp Med Biol. 1992;315:15-22.
 pikfarma.ru
pikfarma.ru