Глутамин – необходимое питательное вещество для больных, получающих интенсивную терапию
СтатьиОпубликовано в журнале:
Int J Colorectal Dis (Международный журнал заболеваний толстой и прямой кишки) »» 1999; 14: 137-142
Ян Вернерман, Фолк Хаммарквист*
Клиника Хаддинг при Каролинском Институте, Стокгольм, Швеция
* J. Wernerman, F. Hammarquist. Dept. of Anesthesiology and Intensive Care and Surgery, Huddinge Hospital, Karolinska Institutet, S-14186 Stockholm, Sweden. Тел. (+46-8) 58586395, факс (+46-8) 7795424, e-mail jan.wernerman@anaesth.hs.sll.se
Резюме.
Глутамин является распространенной аминокислотой, что имеет особое значение для больных, получающих интенсивную терапию. Глутамин может использоваться как субстрат в окислительных процессах, требующих быстрой регуляции в относительно широком диапазоне объемов. У больных, получающих интенсивную терапию, синтез глутамина в скелетных мышцах и его перенос в другие ткани могут быть недостаточными, и таким больных предлагается дополнительно вводить глутамин. Имеются данные о том, что это улучшает долгосрочную выживаемость, что делает применение глутамина одним из немногих способов лечения, улучшающих результаты интенсивной терапии. В данном обзоре описываются метаболические и физиологические свойства глутамина и новейшие клинические данные о его применении.
Введение
Среди 20 аминокислот, присутствующих в организме человека, глутамин (Глн) привлекает особое внимание исследователей в области клинического питания и метаболизма. Хотя глутамин относится к заменимым аминокислотам, он обладает некоторыми свойствами, которые придают ему уникальное значение в условиях усиленного катаболизма. Глн – это самая распространенная свободная аминокислота человеческого организма. Его доля в плазме крови наибольшая среди всех свободных аминокислот (0,5-0,8 из 2,0-2,5 ммоль/л). В мышцах концентрация Глн составляет примерно 20 ммоль/л из общей концентрации всех аминокислот 30 ммоль/л (1). Высокая концентрация Глн (хотя и меньшая, чем в мышцах) наблюдается и в других тканях (слизистой оболочке кишечника, печени, лейкоцитах) (2,3), но в этих тканях некоторые другие заменимые аминокислоты имеют более высокую концентрацию. Из скелетных мышц глутамин выходит в кровь и поступает в кишечник, печень и селезенку (4). Скорость межорганного транспорта всех аминокислот в покое составляет примерно 25 ммоль/час, и треть этого количества приходится на Глн. Большинство аминокислот организма входят в состав белков. Однако Глн не является самым распространенной аминокислотой как элементом белка; таковой является лейцин, на который приходится примерно 10% всех аминокислот в составе белков. Тем не менее, Глн является одной из самых распространенных аминокислот, и в большинстве белков его доля составляет 5-7%.
При первоначальной разработке препаратов для парентерального питания в качестве белкового компонента использовали гидролизат казеина (5). Этот гидролизат содержал свободные аминокислоты и олигопептиды. Такие препараты никогда не были абсолютно стандартными, и иногда олигопептиды оказывались достаточно большими, чтобы вызывать аллергические реакции. В конце 1950-х г.г., когда появились растворы кристаллических аминокислот, Глн в них не включали из-за того, что он имеет низкую растворимость и нестабилен в водном растворе. Глутамин имеет тенденцию к образованию ортоглутамата, который может вызывать некоторые побочные эффекты. Кроме того, регистрация нестабильного препарата контролирующими органоми невозможна. В то время значение Глн с метаболической точки зрения было не полностью понятно. Глн синтезируется в скелетных мышцах и оттуда через кровь попадает в печень и селезенку. В печени от Глн отщепляется аминогруппа и образуется глутамат (Глу), который переносится через кровь (главным образом в эритроцитах (6)) обратно в периферические ткани. В мышцах Глу снова превращается в Глн, и, таким образом, по этому механизму аммоний из периферических тканей переносится в печень.
Метаболизм
Два фермента играют наибольшую роль в метаболизме Глн: глутамин-синтаза и глутаминаза (таблица 1).
Таблица 1.
Наличие глутамин-синтазы и глутаминазы в разных тканях
|
Глутамин-синтетаза |
Глутаминаза |
|
|
Скелетные мышцы |
++ |
+ + + ++ |
Глутамин-синтаза присутствует в высокой концентрации в мышцах, печени и легких. Он катализирует реакцию образования Глн из Глу путем присоединения аминогруппы. Окисление аминокислот в мышцах является наиболее важным в количественном отношении способом переноса аминогрупп в печень для образования мочевины и выведения азота из организма. Альтернативным способом выведения азота является образование аммиака, который может оказывать нейротоксическое действие, и поэтому непригоден для транспорта азота вне печени, которая выполняет функцию фильтра аммиака. Другой возможностью является образование аланина (Ала) из пирувата в цикле Кори. В результате этого Глн и Ала составляют примерно 70% аминокислот, выделяющихся в кровь из периферических тканей в спокойном состоянии (5). В состояниях, сопровождающихся усиленным расщеплением белков мышц, реакции окисления аминокислот и образования аммиака усиливаются, поэтому образование Глн и Ала и выделение в кровь тоже может усиливаются. В таких стрессовых состояниях выделение аминокислот из периферических тканей увеличивается в 3-4 раза, соотношение отдельных аминокислот поддерживается (7). После приема пищи часто наблюдается суммарное накопление аминокислот в мышцах, но, тем не менее, всегда осуществляется выделение Глн и Ала, синтезируемых по описанному выше механизму (8,9).
Глутаминаза наиболее распространена в тканях, захватывающих Глн (10). Слизистая оболочка кишечника, печень и иммунокомпетентные клетки являются примерами тканей со сравнительно небольшим градиентом концентрации свободного Глн между внутри- и внеклеточным пространством. Захватываемый клетками этих тканей глутамин быстро расщепляется глутаминазой до глутамата. Образующийся Глу в этих тканях используется в основном в процессах энергетического метаболизма. При полном окислении Глн может образоваться 30 моль АТФ/моль Глн. Глн используется в качестве энергетического субстрата во многих тканях, и, по крайней мере в экспериментальных клеточных культурах, клетки слизистой оболочки кишечника, печени и иммунокомпетентные клетки используют больше Глн, чем глюкозы (11-13). Использование глутамина как приоритетного источника энергии еще более усиливается при некоторых стрессовых состояниях. В классической работе Виндмюллера и Спэта (14) показано, что в при расщеплении Глн в кишечнике крысы образуется Глу, Ала, лактат и аммиак.
Глн является одним из основных субстратов, используемых для синтеза пуринов и пиримидинов. Наличие Глн является необходимым условием синтеза нуклеотидов. Для обеспечения деления быстроделящихся клеток важно, чтобы синтез нуклеотидов мог быстро усиливаться. Ньоюсхолм с соавт. (15) предположили, что метаболическая регуляция синтеза
нуклеотидов также связана с регуляцией использования Глн в качестве субстрата окислительных процессов. Большой оборот в пути окисления Глн позволяет направить часть Глн на синтез пуринов и пиримидинов. Даже если эта часть будет составлять всего несколько процентов от количества окисляемого Глн, то синтез пуринов и пиримидинов может мгновенно усилиться в 100 и более раз. Хотя эта утверждение метаболической регуляции носит теоретичесткий характер, оно представляется весьма привлекательным и объясняет важную роль Глн в метаболизме нуклеотидов (16).
Физиологические состояния
При кратковременном голодании концентрация Глн в мышцах человека снижается (17), а после восстановления питания она быстро нормализуется (рис.1).
Рис. 1.
Концентрация свободного глутамина в скелетных мышцах здоровых добровольцев (n=8) в процессе 3-дневного голодания снижается, а после 2 дней приема пищи восстанавливается (из работы 17).

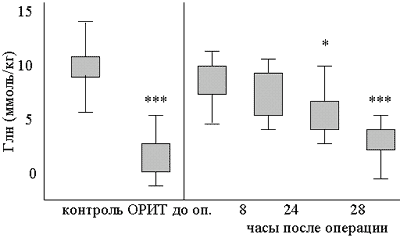
Концентрация Глн в мышцах также снижается после инфузии адреналина или комбинации гормонов стресса в физиологической концентрации (18). Однако в последнем случае концентрация Глн восстанавливается медленно. Более того, снижение концентрации свободного Глн в мышцах продолжается и становится еще более глубоким даже через 18 часов после нормализации уровня гормонов стресса в плазме крови (19). Это свидетельствует о том, что регуляция концентрации Глн в мышцах не связана непосредственно с гормонами стресса, и механизм этой регуляции еще подробно не изучен. У животных обнаружена статистически достоверная корреляция между концентрацией Глн в мышцах, сниженной в результате недостаточного питания или действия эндотоксинов, и синтезом белков в мышцах (20). У людей снижение концентрации свободного Глн в мышцах после хирургических операций статистически достоверно коррелирует со снижением концентрации полирибосом в мышцах (21). Однако глубокое снижение концентрации свободного Глн в мышцах, наблюдаемое у больных, получающих интенсивную терапию, не сопровождается столь же сильным торможением синтеза белков (22,23). Кроме того, концентрация свободного Глн в мышцах нормализуется к 20 дню после операции, а концентрация рибосом в мышцах не восстанавливается и через 30 дней после операции (24,25). Таким образом, иногда наблюдается зависимость между синтезом белков и концентрацией Глн в мышцах. Голодание и хирургические операции не сопровождаются существенным изменением концентрации Глн в слизистой оболочке кишечника (2,26). После операций концентрация Глн в мышцах падает на 40%, а в плазме крови - не изменяется или изменяется незначительно (27). У больных, получающих интенсивную терапию, концентрация свободного Глн в мышцах за несколько дней падает до уровня менее 25% от нормального (рис.2), а в плазме крови – не изменяется или снижается максимум на 30% (22,28). Одновременно усиливается выведение Глн из мышц и захват Глн в селезенке. В настоящее время не ясно, сколько времени сохраняется это усиленное выведение Глн. Однако уменьшение объема мышечной ткани у больных, получающих интенсивную терапию, происходит очень быстро – до 10% в неделю в первый месяц лечения (31,32).
Рис. 2.
Концентрация свободного глутамина в мышцах здоровых добровольцев, больных, получающих интенсивную терапию (n=8+8), и больных, подвергавшихся абдоминальным операциям (n=8). Показана очень низкая концентрация у больных, находящихся в критическом состоянии, и снижение концентрации после операции (из работ 29,30). ОРИТ – больные из отделения реанимации и интенсивной терапии
Метаболизм Глн в органах иммунной системы изучен слабее. Исследования на животных показали, что Глн утилизируется и в слизистой оболочке кишечника, и в иммунокомпетентных клетках (33,34). В кишечнике и селезенке обнаружен захват Глн (35). Исследования метаболизма Глн в органах иммунной системы людей немногочисленны (6,36,37). Во время хирургических операций захват Глн наиболее выражен в тонкой кишке и селезенке (37). Концентрация Глн в плазме крови и слизистых оболочках снижается при недостаточном питании, но после хирургических операциях и в начальных стадиях тяжелых заболеваний концентрация Глн в слизистой оболочке кишечника изменяется лишь незначительно (37,38). Косвенным свидетельством накопления Глн в кишечнике является ослабленный метаболизм Глн у больных, подвергавшихся резекции кишечника (39).
Считается, что при проведении парентерального питания кишечник находится в особо неблагоприятных условиях. Однако лишь в очень немногих контролируемых исследованиях это предположение было подтверждено и были изучены механизмы этого нарушения (40). Перекрестный анализ результатов 27 контролируемых исследований, в которых участвовали больные с желудочно-кишечными, нейрохирургическими, онкологическими и гематологическими заболеваниями и больные, находящиеся в критическом состоянии, не выявил различий объективных показателей при энтеральном и парентеральном питании (41). Авторы сделали вывод об отсутствия влияния пути введения пищи. Тем не менее, при отсутствии энтерального питания наблюдается атрофия слизистой оболочки кишечника, и, как полагают, это может сопровождаться проникновением бактерий через стенку кишечника и развитием сепсиса. Проникновение бактерий через стенку кишечника продемонстрировано в ряде моделей у животных, но патофизиологическое значение этого явления у человека неясно. Здоровые люди способны адаптироваться к парентеральному питанию после некоторого адаптационного периода, и поэтому никаких метаболических различий при парентеральнои и энтеральном питании не обнаруживается (42). Аналогично, больные, подвергавшиеся резекции кишечника, десятилетиями постоянно получают парентеральное питание. С другой стороны, при стрессе ситуация совсем другая, и сейчас появляется все больше данных в пользу преимущества добавления Глн в схемы питания, независимо от пути введения пищи.
Добавление глутамина
Эксперименты на животных четко продемонстрировали важную роль Глн при ряде стрессовых состояний. На животных также показано, что добавление Глн в программу нктриционной терапии оказывает благоприятное действие, особенно на кишечник и иммунную систему. У людей показано благоприятное влияние Глн на морфологию слизистой оболочки кишечника здоровых добровольцев и больных с желудочно-кишечными заболеваниями (43,44). Кроме того, при добавлении Глн наблюдается улучшение всасывания веществ в кишечнике больных с желудочно-кишечными заболеваниями и у больных, находящихся в критическом состоянии (43,45). Сложнее продемонстрировать улучшение функций иммунной системы. Однако известно, что добавление Глн в схему питания послеоперационных больных стимулирует митоз лимфоцитов in vitro (46), а у больных с панкреатитом стимулирует выброс интерлейкина-8 из моноцитов (47).
Глн ослабляет потерю мышечной массы, наблюдающуюся при ряде стрессовых состояний. Большое количество исследователей отмечали улучшение общего баланса азота (48,51). Более того, добавление Глн в схему питания послеоперационных больных способствует поддержанию синтеза белков в мышцах (49) и препятствует снижению концентрации свободного Глн в мышцах (48,49). Влияние на концентрацию Глн в мышцах также наблюдается при применении одного Глн (52), и этот эффект исчезает при прекращении добавления Глн в пищу (53).
У гематологических больных, получающих тотальное парентеральное питание с добавлением Глн, снижена частота инфекций (51,54) и ослабляется внеклеточное накопление воды в процессе химиотерапии (54,55). Также предотвращается развитие вызываемого химиотерапией стоматита (56). Добавление Глн к стандартной программе, изолированно или в сочетании с другими питательными веществами, снижает частоту инфекций у больных с травмами (57-59). Имеется лишь несколько сообщений о влиянии Глн на результат лечения больных. При проведении клинического питания часто встречаются ситуации, когда трудно продемонстрировать улучшение клинического состояния больного (60). Ясно, что нарушение питания сопровождается повышенной заболеваемостью и смертностью (61,62), и профилактика нарушений питания является важной задачей лечения. Профилактика инфекций у больных, подвергавшихся пересадке костного мозга, сокращала только длительность пребывания в клинике (51). Однако если профилактика недостаточности питания является основной целью, то различие смертности может быть выявлено при включении в анализ не только периода пребывания в клинике, но и периода восстановления. В хорошо спланированном рандомизированном контролируемом исследовании с участием больных, получающих интенсивную терапию, Гриффитс с соавт. (63) показали снижение смертности больных при сроке наблюдения 6 месяцев. Это единственное исследование, в котором обнаружено различие результата лечения, но в нем участвовали менее 100 больных из одной клиники. Этот результат еще нужно подтвердить, но на данный момент это исследование является сильным аргументом в пользу добавления Глн в схемы долгосрочного парентерального питания больных, получающих интенсивную терапию.
Рекомендации по питанию
Энтеральное питание рекомендуется всем больным, у которых имеется и функционирует желудочно-кишечный тракт. Эффекты добавления Глн в схемы энтерального питания изучены значительно хуже, чем эффекты добавления Глн в схемы парентерального питания. В описанных выше исследованиях с участием больных с травмами показано влияние Глн на развитие ряда инфекций (57-59), но клиническую значимость этих результатов еще предстоит установить.
Более надежно описаны эффекты добавления Глн в схемы парентерального питания, проводимого изолированного или в сочетании с энтеральным питанием, особенно у гематологических больных и больных, получающих интенсивную терапию (51,54,63). Сведения о дозах добавляемого Глн противоречивы, и об этом требуется дополнительная информация. Больные с гематологическими заболеваниями получали Глн в дозе 40 г/сут (51,53), а больные, получающие интенсивную терапию – 20-25 г/сут (61). В исследованиях с участием больных с желудочно-кишечными заболеваниями, у которых было выявлено влияние Глн на морфологию слизистой оболочки, применялась доза Глн 12,5 г/сут (43); влияние на мышечную ткань показано при дозе Глн 20 г/сут (48,49,50). Исследования дозовой зависимости эффектов Глн до сих пор не проводились. Поэтому основанием для добавления Глн в схемы питания является в основном обеспечение проведения полного питания, включающего все необходимые питательные вещества. Однако полное энтеральное питание недостаточно для поддержания концентрации Глн и синтеза белков в мышцах больных, подвергавшихся хирургическим операциям (53,64).
В большинстве исследований использовались разные экспериментальные препараты, в том числе раствор L-Глн в воде (65). Однако, поскольку L-Глн нестабилен в воде, такой препарат невозможно производить в коммерческих масштабах. Решением этой проблемы является внедрение дипептидов глицил-глутамин и аланил-глутамин (66). После инфузии здоровым добровольцам эти дипептиды быстро расщеплялись; то же наблюдается у больных с нарушениями функций печени или почек (67,68). Существует 2 основных способа добавления Глн в растворы аминокислот для парентерального применения: путем составления совершенно новой смеси, содержащей Глн (69), или путем создания добавки, которую смешивают с имеющимся раствором. Первый путь, конечно, удобен и экономичен, но в некоторых случаях фиксированное соотношение общего количества азота и количества Глн в смеси не обеспечивает достаточной гибкости применения препарата.
Литература
1. Bergstrom 1, Furst, et al (1974) Intrace11ular free amino acid concentrations in human muscle tissue. J Appl Physiol 36;693-697
2, Ahluman et al (1994) Short term starvation alters the free amino acid content of human jntestina1 mucosa. Clin Sci 86:653-662
3. Barle H et al (1996) The concentrations of free amino acids in human liver tissue obtained during laparoscopic surgery. C1in Pbysiol 16:217-227
4. Felig et al (1971) Amino-acid metabolism in exercising man. J ÑÈï Invest 50:2703-2714
5. Elman R (1936) Òhe time factor in retention of nitrogen after intravenous injection of mixture of amino acids. ðòå Soc Exper Biol Med 40:484-487
6. Felig et al (1973) Evidence of inter-organ aøi no-acid transport Üó blood cells in Üwnans. ðòå Natl Àñàä Sci USA 70:1775-1779
7. Clowes et al (1980) Amiïî acid and energy metabolism in çåðéñ and traumatized patients. J Parent Enler Nutr 4: 19S-203
8. Elia et al (1983) Effects of ingested steak anä infused leucine îï forelimb metabolism in òàï and tbc fate of ñÜå carbon skeletons and aøiïî groups of branched-chain aøiïî acids. Clin Sci 64:517-526
9. Wahren J, et al (1976) Effect î! protein ingestion îï splanchnic ànd leg metabolism in ïînnai òàï and patients witb diabetes mellitus. J ÑÈï Invest 57:987-999
10. Meisler (1984) Enzymology of glutamine. In: H:lUssinger D. Sies Í (eds) G1utamine metabolism in mammalian tissue. Springer, BerHn Heidelberg New York, ðð 3-15
11. Souba WW. Smith Þ. Wilmore DW (1985) Glutamine metabolism in the intestinal tract. J Parent Enter Nutr 9:608-611 12. Haussinger D (1989) glutamine metabolism in the liver: îóè view àïdñuïåït conceplS. Melabolism 38 [Suppl]:I4-17
13. Calder (1995) Glutamine action in cells of tbe immune system. In: Cynober L, FUrst Ð, Lawin Ð (edç) Pbarmaceutical nutrition. Immune nutrition. Zuckschwerdt. Munlch, ðð 1020
14. Windmueller et al (1974) Uptake and metabolism of plasma glutamine Üó tbe small intestine. J 8101 ÑÜåò 249: 5070-5079
15. Newsholme et al (1985) Glutamine metabolism in Iymphocytes: 118 ûochemical, pbysiological and clinical importance. Q J Åõð Physiol 70:473-489
16. Souba WW et al (1990) Òhe role of glutamine in maintaining à Üåàløó gï. and supporting øå metabolic response 10 jnjury and infection. J Surg Res 48:383-391
17. Andersson et al (1994) The effcet offasting îï muscle gllltathion 1eve\s. Clin Nulf 13 [Supp1]:12
18. Wemerman et al (1993) Slfess hormones alter (Üñ ðàÍåò ot"I'Tce amino acids in human skeletal muscle, Clin Physiol 13 :309-3 19
19. Ejesson et al (1990) Stress Üîãmones initiate changes in mu$clc [.'åå amino ncid concentTaüîns eharacteTistie fOT suTgical tTaumn, C\in Nutã 12[Suppl]: 39
20. Jepson et al (1988) Re\ationship between glutalnillC concentTation and ðòîtein synthesis in ñà! skeletal mllscle. Àm J Physio\ 255: Å166-®172
21. Wemerman et al (1990) A1pha-ketoglutarate and postoperalive ïþsñlå catabolism. Lancet 335: 701-703
22, Roth et al (1982) Mcl<1l)()lic disordel"s in seveTe àÜdominal sepsis: glutamine dñÏñiñïñó min ske\eta1 musc\e. Ñliï NutT 1:25-42
23. Essen et al (1998) Tissue prolein synthesis in the eritical1y ilI patient, Cril Ñàl"å Med 26:92-100
24. Petersson et al (1990) Elective nbdominal suTgery depresses muså1å prolein synthesis and increases subjective fatigue for òîãåtha" 30 dnys. ®ñ J Surg 77:796-800
25, Pelersson et al (1992) Long-term changes in muscle Il'ee amino acid levels àÑlåã å1åñèóå abdomina1 surgery. ®ã J SUI'g 79:212-216
26. Ahlman et al (1995) E1ective abdominal operations a1ter [Üå free àIÏiïî acid eOnlent of the hum:ø intestinal mucosa. Åèñ J Surg 161:595-603
27. Vtnnars et al (1975) Influence of (Üå post operative stale îï the intTaceHllla.' fTee amino acids in human muscle tissue. Àïï Surg 182:665-671
28 Gamin L et al (1996) À descriptive study 01' skeletal musc1e metabolism in eritica1ly i1l patients: free nmino acids, energy-I'ich phosphates, protein, nucleic acids. fat. watcr, alld e\ectrolyles, Crit Ñàãå Med 24:575-583
29, Hammarqvisl et al (1997) Skelctal JlIuscle glulaJhione is depleted in critica11y iII patients. Ñï! Ñàãå Ìñè 25:78-84
30. Luo J-L. et al (1998) Surgical trauma decreascs tllC ynlJlesis of glulathione in skcleta1 musc1e, Àò J PhysioI275:E359-Å365
31. Streat et al (1987) Aggressive nutritiona1 suppon does not prevent protcin los dcspile 1'lIt gain io septic
ioteosive eare patleots, J Traull1a 27:262-266
32. Gamrin et al (1997) Longi!Utlil1al ñløïgås îå biochemica1 ðàrameterS in muscle during c.'ili..:al iI1ness. MelaboBsm 46: 756-762
33. WindmuelIer et al (1980) Respiratory fields and nitrogen metabo1ism in vivo in small intestine of fed rats. Quantitative importance of glutall1ine. glutønàlå and aspartate. J 8io1 ÑÜåm 255:107-112
34. Newsho1me et al (1989) Glutamine metabolism in different tissues: its physiological nod pathological\ importance. 10: Êinnåó JM, BO1ilfi1"PR'(eds) Verspectivcs ill clinica1 l1utrition. ØÜnï & Schwarzenberg. 8à1üòîãå. ðð 71-97
35. Deutz et al (1992) Postoperative changes in hepatic inLcstinal. splcnic and muscle fluxes of arïiïî acids al1d øønîïin in pigs. Ñ®ï Sci 83: 607-614
36 McAena et al (1991) Selective uptake î! glUlàùiïñ in (Üå gastrointestina1 tract: confirmalion in à humnn slutly. ®ã J Surg 78:480-482
37. Hu1st et al (1997) Glutamine extraction Üó the glut and Jhe effect of nutritiona1 dep1etion. Ànn Surg 225:112-121
38. Ahlman et al (1995) lntestinal arïi"î acid contcnt in critiea11y iII patients. J Parenl Bnler Nutr 19:272-278
39. Darmaun et al (1991) Glutamine metabolism afler SmaII intcstina1 resection in hwnans. Metabolism 40:42-44
40. Buchman et al (1995) Pareotera1 nutrition is associated with intestina1 morpho1ogic anä funetiona1 ñÜangåà in Üîòans. J Ðçråï' Enler Nutr 19:453-460
41. Dunham Ì (1997) Enlema1 nutrition does ïî! decreasc sirs, mods and mortality. Shock 7 [Supp11]:147
42, Khan et al (1992) Nutrient utilisation in mus ñ1å and in Jhe whole body ofpatients receiving tota1 parentera1 nutrition. ÑÍï Nutã 11:345-351
43. Hulst et al (1993) ÒÜå role of parentera1 glutamine administration in preserving gul integrity. Lancet 334:1363-1365
44. Scheppach et al (1994) Øfåñ! of free g1utamine and a1anyl-glutamine dipeptide îï mucosal prolifera. tion of the Üwnan i1åwn and åî10Ï. Gastroenterology 107: 429-434
45. Treme1 et al (1994) Glutamine dipeptide-supplemented parentera1 nutrition øaiï. tains intesti"a1 Ñîïñèî" in the critica11y Ø. Gastroentero10gy 107:1595-1601
46. O'Riordan et al (1994) GJutamine supp!emented tota1 parentera1 nutrition ånhanñå! T-1ymphocyte ãå®ðîï®å in surgica1 patients undergoing colorecta1 resection. Àïï Surg 220:212-221
47. Beaux et al (1998) Glutamine-supp1emented tota1 parenteraI nutrition reduces blood mononuc1ear ñå11 inter1eukin-8 release in severe pancreatitis. Nutrition 14:261-265
48, Stehle et al (1989) Effect of \larenteral g\utamine peptide suoo1emenls îï musele g!utam1ne 10ss and nitro®å" balanñå after major surgery. Lanñå! 1:231-233
49. Hammarqvist et al (1989) Addition of glutamine to total parenteral ïîtrition afler e1ective abdomina1 surgery sparea free glutamine in musc1e. counteraets the fa11 in muscle prolein synthesis. and improves nitrogen balancc. Ànn 5urg 209:455-461
50. Hammarqvist et al (1990) Alanyl-glutamine counteracts the dep1etion offree glutøniïå and the postoperative äåñliïå in protein synthesis in ske1eta1 muscle. Àïï Surg 212:637-644
51. Ziegler et al (1992) Clinica1 and metabolic åfáñàñó of g1utamine.supplemented parentera1 nutrition after Üîïå marrow transplantation. Àïï 1ntern Ìåä 116:821-828
52. Blomqvist et al (1995) 01utamine and a1pha-ketog1utarate ðãåóåï! the decrease in muscle free glutamine eoneentration and inf1uence protein synJhesis after total Ûð replacemenL Metabo1ism 44:1215';'1222 –
53. Petersson et al (1994) Long-term effect of glyey1-g1utamine after elective surgery îï free amino acids in muscle, J Parent Enter Nutr 18:320325
54, Sch1oerb et al (1993) Tota1 parenteral nutrition wiJh glutamioc io Üîîå marrow transp1antation and other ñliniñà1 àððliåàÜÎnS (à randomized double-blind study). J Ðàòåï! Enter Nutr 17:407-413 142
55. Scheltinga et al (1991) G1utamiëå-ånriñhåd intravCnOHs I'cedings àttñïíàlå extracellular fluid åõðàïsiîë after à ,tàùJàãd stress, Àëï Surg 214: 385-395
56. Skubilz et al ( 1996) Îãàl gl utàmiëå 10 ðãåvåët chemotherapy induced stomatitis: à pilot stttdy, J Lab Ñliï Ìåä 127:223-228
57. Moore et al (1994) Ñliëiñàl Bebefits of àï Immuëå-åëhàïñiëg dlet for ca]'ly postinjury cnteraJ feeding. J Trauma 37:607-615
58. Kudsk et al (1996) À l'andomized traH of isîëitãîgåïîus enleral dicts after sevcre trauma - àë immune.enhancing diet reduces septic complications, Àëï 5urg 224:531-540
59. Houdijk et al (1998) Glutamine ñïriched enteral nutrition ãåäèññ,; i/1I'ectious mîãÛditó in multitrauma patients: à doubIe-bIiïd, pl'ospective, ãàïdOlnizåd trial. Lancet 352:772-776
60. Souba (1997) How should wc cvaluate the el't'icacy î! ëu trition support? J Òãàèòà 42:343-344
61. Giner et al (1996) À ñîããålalion between malnutrition Ùld ðîîã outcome in ñãiticàlló-iII patients sti1l exists. Nutãltiîï 12:23-29
62. McWhirter JP, Pennington CR (1994) Incidence and recognition î! malnutrition in hospitaJ. ®Ì] 308:945
63. Griffith RD et al (1997) À six.month outîòå of critica1Jy iI1 patients given glutamine supplemented parenteral nutrition. Nutrition 13:295-302
64. Petersson et al J (1994) Long-term effect î! postoperative ÒPN supplemented with glycyl.glutamine îï subjective fatigue and muscle ðro. tein synthesis. ®ã J Surg 81:1520-1523
65. Khan et al (1991) Òhe stability î! L-glutamine ië total parenteral nutrition solutions. Clin Nutr 10:193-198
66. Furst et al (1990) Dipeptides in ñliniñà1 nutrition, Ðãîñ Nutr ®îñ 49:343-359
67, Albers et al (1988) ÀvàilàÜØtó of àrïiïî acids supplied intravenously in healthy òàï as synthetic dipeptides. Pharrnacokinetic evaluation of Làlàïól-L-glutamiëå and L-glóñól-L-glóñól-IÀyrosiïå. Ñliï Sci 75:463-468
68. Hubl et al (1994) importance of lióåã and kidney function {îã the utilization of glutamine-containing dipeptides i/1 òan. Metabolism43:1104-1107
69. Wiren et al (1995) The effect of à new àmiëî-àñid dipeplide solution îï protein metabolism and humoral growth factors in the postoperative state in òà/1, Ñliï Nutr 14:97-104



