Амиксин: индукция интерферонов-α, -β, -γ И -λ в сыворотке крови и легочной ткани
СтатьиС. Григорян1, доктор медицинских наук, профессор, Е. Исаева1, кандидат биологических наук, B. Бакалов1, Е. Осипова2, кандидат медицинских наук, А. Бевз2, кандидат медицинских наук, И. Простяков3, кандидат медицинских наук, C. Надоров3, кандидат биологических наук
1Федеральный научно-исследовательский центр эпидемиологии и микробиологии им. Н.Ф. Гамалеи, Москва
2ПАО Отисифарм, Москва
3ОАО Фармстандарт, Долгопрудный E-mail: aybevz@otcpharm.ru
В строго контролируемых лабораторных условиях, соответствующих принципам доказательной медицины, идентифицированы все известные 3 типа ИФН (α/β, γ,λ), которые индуцируются препаратом Амиксин в крови и легочной ткани. Подробно изучены динамика, последовательность и профиль их системной и локальной продукции. Индукция препаратом Амиксин эндогенных ИФН 1-го, 2-го и 3-го типов в крови и легочной ткани активирует врожденный и адаптивный иммунный ответ, сохраняет иммунный гомеостаз собственного организма и предотвращает и (или) прерывает синтез вирусных нуклеиновых кислот и вирус-специфических белков, не вызывая характерных для экзогенных препаратов ИФН побочных эффектов.
Ключевые слова: индукция синтеза интерферонов, Амиксин, активация врожденного и адаптивного иммунного ответа.
Амиксин (тилорон) — низкомолекулярный синтетический индуктор синтеза интерферона (ИФН) — относится к классу флуоренонов, ИФН-индуцирующая способность которого при пероральном введении была впервые установлена в 1970 г. G. Mayer, R. Kruger [1]. Из имеющихся сегодня на фармацевтическом рынке индукторов ИФН Амиксин наиболее изучен. Более чем 30-летний опыт клинического применения свидетельствует о его безопасности и эффективности при широком круге заболеваний инфекционного и неинфекционного генеза [2]. Несмотря на разнообразие генетического материала вирусов, Амиксин, как и ИФН, подавляет фундаментальные процессы их репродукции на стадиях, обязательных для всех вирусов: блокирует адсорбцию вирусов; синтез вирусспецифических белков и сборку зрелых вирусных частиц, распознавая и дискриминируя вирусные информационные РНК от клеточных; подавляет внутриклеточное размножение вирусов. Именно поэтому к действию Амиксина чувствительны практически все РНК- или ДНК-содержащие вирусы [3, 4].
Семейство ИФН является ключевым компонентом врожденного и адаптивного иммунитета, обеспечивающим 1-ю линию защиты организма от инфекционных агентов [5]. В настоящее время идентифицированы 3 типа ИФН:
- ИФН 1-го типа, включающий в себя прежде всего ИФНα/β, который в той или иной степени продуцируется всеми ядерными клетками, оказывает прямое противовирусное действие на репликацию вирусов в инфицированных клетках, предупреждает инфицирование окружающих и, активируя каскад антивирусных сигнальных путей, включает врожденный иммунитет и способствует развитию соответствующих адаптивных иммунных ответов [6, 7];
- ИФН 2-го типа — ИФН-γ, который продуцируется разными субпопуляциями Т-лимфоцитов и естественных киллеров — NK, регулируя гомеостаз в лимфатических узлах и очагах инфекции, вызывает стимуляцию и обеспечивает функциональную эффективность специфического адаптивного иммунитета [8];
- недавно открытый 3-й тип ИФН (ИФНλ) представлен ИФН λ, - λ2, - λ3 (известными также как интерлейкин — ИЛ — 29, -28A и -28B), функционально тесно связан с ИФН 1-го типа, экспрессия рецепторов которого, однако, определяется на узком спектре клеток, ограничивая ответ на ИФН 3-го типа [9]. Мишенью ИФНλ, являются прежде всего эпителиальные клетки дыхательных путей и легких, желудочно-кишечного тракта (ЖКТ), половых органов, кожи, гепатоциты печени, мононуклеарные клетки периферической крови — peripheral mononuclear blood cells (РМВС) и плазмоцитарные дендритные клетки [10—13]. Антивирусное действие ИФН 3-го типа дополняет антивирусное действие ИФН 1-го типа; более того, оно преобладает над таковым во «входных воротах» инфекции и не вызывает характерных для ИФН 1-го типа системных побочных эффектов [14]. Все 3 типа ИФН выполняют роль 1-й линии защиты организма не только от вирусных, но и от бактериальных и паразитарных заболеваний [15, 16]. Важно отметить, что ИФН продуцируются не только при разных видах инфекционной патологии, но и под действием ряда высоко- и низкомолекулярных соединений — индукторов ИФН [3, 4].
Известно, что Амиксин стимулирует выработку в организме эндогенных ИФН, которые в ранних исследованиях тестировались биологическим методом — по противовирусному действию — и были отнесены к ИФНα, -β (1-й тип) и -γ (2-й тип) [2, 3]. Хотя 1-й мишенью контакта и воздействия Амик-сина является ЖКТ, где ведущую роль в продукции ИФН выполняют ассоциированные с тонким кишечником лимфо-идные образования (GALT — gut associated lymphoid tissue) — пейеровы бляшки, иммунокомпетентные клетки кишечника обладают уникальной способностью к миграции, и потому их стимуляция взывает активацию иммунной системы не только ЖКТ, но и легочного, урогенитального трактов и системного иммунитета [17]. Амиксин вызывает длительную циркуляцию в крови терапевтических доз ИФН, которые предотвращают инфицирование незараженных клеток и создают барьерное антивирусное состояние. Максимальная продукция ИФН в крови людей после 4-кратного еженедельного приема Амик-сина достигает в среднем 64—128 ед/мл, и она сохраняется до 8-й недели включительно [18].
Для оптимизации клинического применения индуктора ИФН — Амиксина — представлялось целесообразным: в рамках доклинического исследования, в строго контролируемых лабораторных условиях, основанных на принципах доказательной медицины, идентифицировать и провести сравнительное изучение индукции препаратом Амиксин ИФНα (1-й тип), -β (1-й тип), -γ (2-й тип) и -γ (3-й тип) в сыворотке крови и легочной ткани экспериментальных животных с использованием современных методов определения типовой принадлежности индуцируемых ИФН; определить последовательность их продукции, продолжительность аккумуляции и установить зависимость динамики индукции — продукции ИФН 1-го, 2-го и 3-го типов — в сыворотке и легочной ткани от дозы вводимого препарата. Подобные исследования препарата Амиксин ранее не проводились.
Целью настоящего исследования было идентифицировать на молекулярном уровне экспериментальные данные по индукции ИФНа и β (1-й тип), -γ (2-й тип) и -λ (3-й тип) в сыворотке крови и легочной ткани мышей препаратом Амиксин (таблетки, покрытые пленочной оболочкой, 125 мг, производитель ОАО Фармстандарт-Томскхимфарм, Россия) при его однократном пероральном (внутрижелудочном) введении в 3 дозах: 20; 40 и 400 мг/кг, рассчитанных эквивалентно разовой, максимальной суточной и десятикратной суточной дозам для человека [19], и оценить динамику продукции ИФНа, -β, -γ и -λ2/3 в сыворотке и легочной ткани мышей под действием препарата Амиксин в 3 разных дозах в течение 120 ч; продукция ИФН отслеживалась в сроки: 0; 4; 8; 12; 24; 48; 72; 96 и 120 ч.
Исследования проводились в 2 этапа:
- 1-й этап осуществлялся в Институте органического синтеза Латвии (Рига); исследовали динамику индукции и продукции ИФНа, β, -γ и -λ препаратом Амик-син в дозах 40 и 400 мг/кг в сыворотке крови мышей в течение 120 ч;
- 2-й этап осуществлялся в лаборатории индукторов интерферона Федерального научно-исследовательского центра эпидемиологии и микробиологии им. Н.Ф. Гамалеи Минздрава России (Москва); исследовали динамику индукции и продукции ИФНа и -X препаратом Амиксин в дозах 20; 40 и 400 мг/кг в легочной ткани мышей в течение 120 ч.
На 1-м этапе использовались самки 55 мышей линии СВА/СаО1аНМ массой 19—30 г, полученные из вивария Harlan Laboratories (Нидерланды), на 2-м — самки 85 мышей линии СВА/CaOlaHsd массой 18—20 г, полученные из питомника Андреевка Научного центра биомедицинских технологий РАМН.
Содержание животных, их питание и уход за ними в ходе эксперимента соответствовали требованиям Правил проведения работ с использованием экспериментальных животных (Приказ Минздравсоцразвития России от 23.08.10 №708н). Этические принципы обращения с лабораторными животными соблюдались в соответствии с European Convention for the Protection of Vertebral Animals Used for Experimental and Other Scientific Purposes. CETS №123. Перед исследованием животные подвергались 5-дневному карантину
Исследуемый препарат — Амиксин (тилорон), лекарственная форма: таблетки, покрытые пленочной оболочкой, 125 мг, ОАО Фармстандарт-Томскхимфарм, 634009 (Россия, Томск).
Исследуемый препарат растворяли в стерильной дистиллированной воде в концентрациях, соответствующих вводимым дозам (20; 40 и 400 мг/кг). Экспериментальные животные распределялись на 2 или 3 группы соответственно этапу исследования и дозам вводимого препарата. Подбор животных в группы осуществлялся случайным образом. Каждую дозу препарата Амиксин вводили мышам однократно перо-рально (внутрижелудочно) в объеме 50 мкл специальными дозированными зондами, размеры которых соответствовали физиологической длине пищевода животных. В каждый срок исследования использовали по 3 мыши.
Мышей выдерживали под легким эфирным наркозом до исчезновения ноцицептивных рефлексов. Забор крови производили методом декапитации в стерильных условиях. Образцы крови, полученные в каждый срок исследования, состояли из пула (смеси) крови от 3 мышей в объеме 3 мл (по 1 мл от каждой особи), из которого после центрифугирования в режиме 5000 об/мин в течение 15 мин отбирали надосадочную сыворотку в объеме 1,0 мл в стерильные пробирки типа Эппен-дорф. Сыворотки крови мышей, соответствующие каждому сроку забора, разделяли на аликвоты по 0,3 мл в 3 стерильные пробирки Эппендорф, каждую из которых хранили при температуре -70°C до проведения соответствующего иммунофер-ментного анализа (ИФА). Для получения суспензии легочной ткани обескровленных мышей фиксировали на специальных парафиновых подложках, в асептических условиях вскрывали грудную клетку, изолировали и извлекали легкие (правое и левое), которые последовательно 3-кратно промывали в буфере Hepes и в питательной среде №199, взвешивали и размельчали сначала в ступке, а затем — в специальном гомогенизаторе. Все процедуры проводились в кратчайшие сроки и на холоде. Разрушение клеток ткани легкого достигалось 3-кратным замораживанием и оттаиванием. Готовилась 10% суспензия го-могената в питательной среде №199. Супернатанты суспензии гомогената легочной ткани после низкоскоростного центрифугирования (при 1600 об/мин в течение 30 мин) отбирали и замораживали при температуре -70°С до последующего исследования методом ИФА.
ОПРЕДЕЛЕНИЕ ИФНа, β, -γ И -λ В СЫВОРОТКЕ КРОВИ И ЛЕГОЧНОЙ ТКАНИ МЫШЕЙ
Содержание ИФНа, β, -γ, и -λ2/3 в пробах сывороток и супернатантов 10% суспензии гомогенатов легочной ткани мышей определяли в сроки 0; 4; 8; 12; 24; 48; 72; 96 и 120 ч (3 мыши в каждый срок) после однократного перорально-го (внутрижелудочного) введения Амиксина в дозах 20; 40 и 400 мг/кг.
Содержание ИФНа, β, -γ и -Я2/3 (ИЛ28А/В) в сыворотке крови и надосадочной жидкости 10% суспензии легочной ткани c последующим пересчетом соответственно на 1 мл и 1 мг легочной ткани определяли методом ИФА с использованием следующих коммерческих тест-систем согласно инструкции производителя:
- Mouse Interferon Alpha ELISA Kit; R&D Systems (США);
- Mouse IFN-p ELISA Kit; PBL Interferon Source (США);
- Mouse IFN-y ELISA Kit; R&D Systems Europe LTD (Великобритания);
- DuoSet® ELISA; Mouse IL-28A/B (IFN-A.2/3) Kit; R&D Systems (CIUA).
Расчет оптической плотности исследуемых проб проводили на спектрофотометре — ридере Anthos Labtec — c использованием вошера и шейкера (Instruments GmbH, Австрия) с последующей обработкой и преобразованием данных в пикограммы в 1 мл и 1 мг (пкг/мл и пкг/мг) по компьютерной программе Software ADAP. Все исследования повторяли 2 раза.
Результаты подвергали статистической обработке путем расчета среднего арифметического (M) и стандартной ошибки (+a). Статистическую значимость различий при межгрупповых сравнениях оценивали, используя двусторонний t-критерий Стьюдента для независимых групп. Для статистических расчетов применяли компьютерную программу Microsoft Excel 2009.
ИНДУКЦИЯ ИФНа, β, -γ И -λ В СЫВОРОТКЕ КРОВИ МЫШЕЙ ПРЕПАРАТОМ АМИКСИН
Однократное пероральное введение Амиксина в дозах 40 и 400 мг/кг вызывало дозозависимую индукцию и продукцию ИФНа и ИФНλ2/3 в сыворотке крови мышей; максимальное их содержание через 24 ч после введения препарата в дозе 40 мг/кг составляло соответственно 176,0±1,0 и 398±12 пкг/мл, а в дозе 400 мг/кг — соответственно 1337+93 и 2231+93 пкг/мл. Уровень продукции ИФНλ2/3 в сыворотке крови мышей под действием Амиксина в дозе 40 и 400 мг/кг в этот срок значительно (соответственно в 7,5 и 5,6 раза) был выше, чем ИФН (рис. 1).
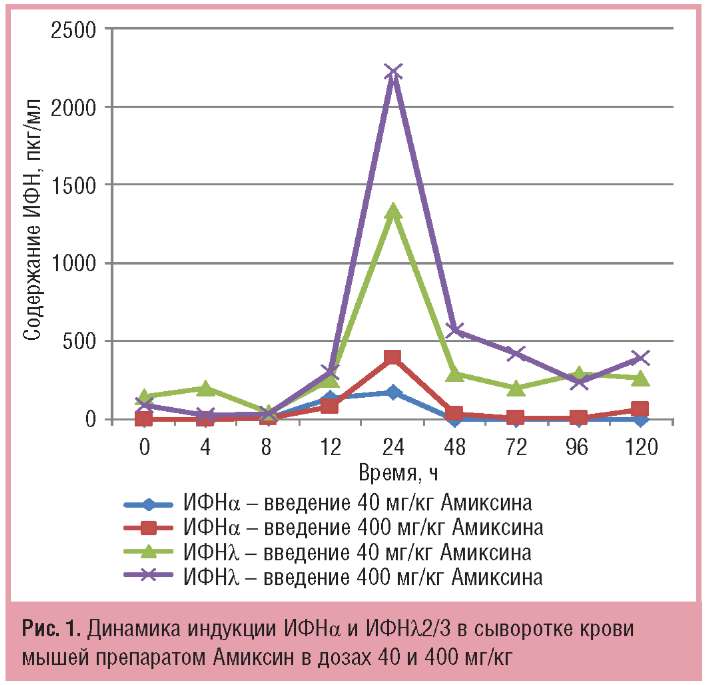
Сопоставимыми были динамика индукции и накопления в крови ИФНβ и -γ под действием Амиксина, максимальный уровень которых достигался также через 24 ч после однократного введения препарата. Однако содержание ИФН-β и -γ в сыворотке крови экспериментальных животных на пике их продукции было ниже, чем ИФН-а и -λ2/3. При дозе Амиксина 40 мг/кг максимальный уровень синтеза ИФН-β и ИФН-γ в крови определялся на уровне фоновых терапевтических значений (соответственно 8,1+4,3 и 4,4+1,2 пкг/мл). При дозе 400 мг/кг максимальные количества ИФНр и -γ через 24 ч после введения препарата увеличивались в среднем в 10 раз в сравнении с таковыми при дозе 40 мг/кг, а в сравнении с контролем — соответственно в 110 и 46 раз (рис. 2).
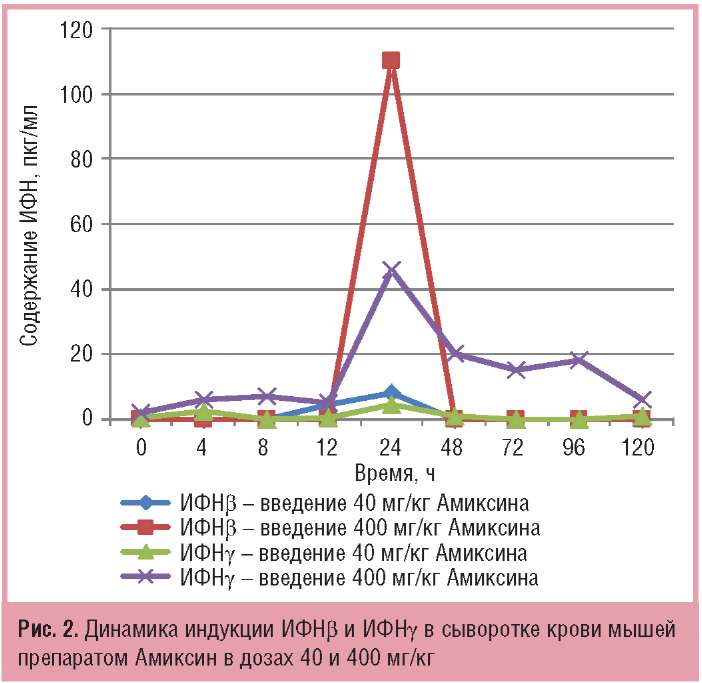
По совокупности результатов, представленных на рис. 1 и 2, однократное пероральное введение Амиксина в дозах 40 и 400 мг/кг вызывало индукцию в сыворотке крови экспериментальных животных всех типов ИФН — ИФНа, β, -γ, -λ, но количество ИФНλ, и α было выше, чем ИФН-β и -γ в среднем в 10 раз.
ИНДУКЦИЯ ИФНа И -X В ЛЕГОЧНОЙ ТКАНИ МЫШЕЙ ПРЕПАРАТОМ АМИКСИН
Таким образом, по результатам 1-го этапа исследования были установлены индукция Амиксином всех 3 типов ИФН в сыворотке крови мышей и преобладание в ней продукции ИФНа и -λ2/3. На 2-м этапе была более детально изучена динамика локальной продукции ИФНа и -λ2/3 в легочной ткани мышей при введении препарата в 3 дозах: 20; 40 и 400 мг/кг.
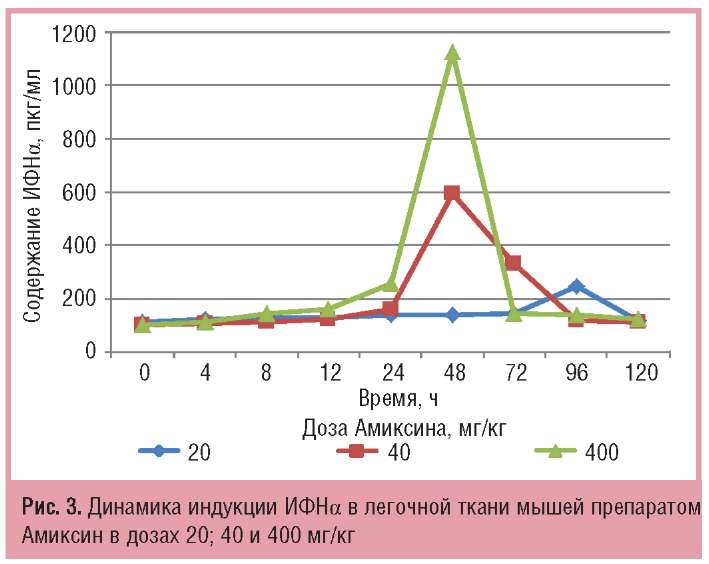
Инициация индукции ИФНа в легочной ткани мышей выражалась в 2-кратном повышении через 24 ч после введения Амиксина в дозе 400 мг/кг среднестатистических значений его выработки относительно нулевой точки, а максимальные количества (1125 пкг/мг) синтезировались и аккумулировались только через 48 ч после индукции той же дозой препарата (рис. 3).
Низкая доза препарата через 96 ч после его введения также вызывала 2-кратное повышение содержания ИФН-а по сравнению с контролем. Увеличение дозы вводимого препарата до 40 мг/кг сопровождалось сокращением срока индукции максимальных количеств ИФНа в легочной ткани мышей к 48 ч (595,0 пкг/мг), причем в течение последующих 24 ч синтез ИФН-а сохранялся на уровне 335,0 пкг/мг
В целом под действием разных доз Амиксина в легочной ткани мышей определялась дозозависимая, но более поздняя, чем в сыворотке крови, продукция ИФН-а (см. рис. 3).
Амиксин в возрастающих от 20 до 400 мг/кг дозах вызывал также индукцию продукции в легочной ткани мышей ИФНλ, среднестатистические значения количества которого на пике продукции составляли соответственно 675,0 и 1547,5 пкг/мг.
Из данных, представленных на рис. 4, следует, что через 48 ч после введения Амиксина в низкой дозе — 20 мг/кг — в легочной ткани мышей определялось 6-кратное повышение содержания ИФНλ2/3 относительно контроля в нулевой точке.
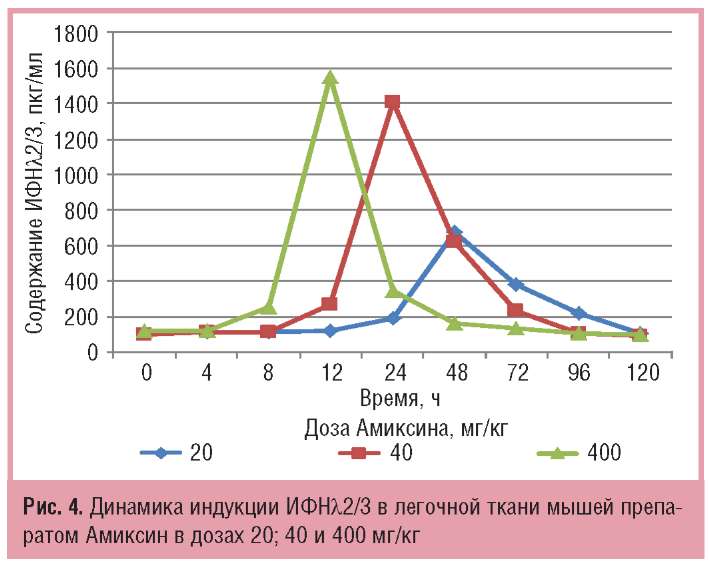
Увеличение дозы препарата до 40 мг/кг повышало содержание в легочной ткани мышей ИФНλ2/3 до 1405,0 пкг/мг и сокращало срок индукции его максимальной продукции на 24 ч относительно низкой дозы. Иными словами, 2-кратное увеличение дозы однократно вводимого Амиксина приводило к 2-кратному повышению количества синтезированного ИФНλ2/3 и 2-кратному сокращению срока достижения пика его продукции. Максимальная доза препарата — 400 мг/кг — смещала пик индукции ИФНλ2/3 в легочной ткани мышей к 12 ч, а уровень его продукции достигал 1547,5 пкг/мг. При использовании Амиксина в дозах 20 и 40 мг/кг продолжительность синтеза ИФНλ2/3 относительно нулевой точки и его аккумуляции в легочной ткани экспериментальных животных составляла соответственно 96 ч и 72 ч. Максимальная доза препарата — 400 мг/мг — вызывала более быстрый (12 ч) и значительный подъем продукции ИФНλ2/3 (1547,5 пкг/мг), которая сохранялась на достаточном уровне (347,5 пкг/мг) относительно контроля до 24 ч включительно.
Таким образом, динамика продукции ИФНλ2/3 в легочной ткани мышей показала, что увеличение однократной пероральной дозы препарата Амиксин сокращает срок достижения пика продукции ИФНλ2/3. В дозах 20; 40 и 400 мг/ кг максимальный синтез ИФНλ2/3 в легочной ткани тестируется соответственно через 48; 24 и 12 ч после введения препарата. При этом 2-кратное повышение дозы Амиксина с 20 до 40 мг/кг вызывает более чем 2-кратное увеличение количества синтезированного в легочной ткани ИФНλ2/3 на пике его максимальной продукции соответственно с 675,00+5,94 до 1405,00+9,86 пкг/мг. При последующем 10-кратном повышении дозы препарата — с 40 до 400 мг/кг — срок достижения максимальной продукции сокращается до 12 ч, но ее уровень практически сохраняется таким, как при введении препарата в дозе 40 мг/кг через 24 ч — соответственно 1547,5+3,7 и 1405,00+9,86 пкг/мг (см. рис. 4).
СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ДИНАМИКИ ПРОДУКЦИИ ИФНа И -λ В СЫВОРОТКЕ И ЛЕГОЧНОЙ ТКАНИ МЫШЕЙ ПРЕПАРАТОМ АМИКСИН
Сравнительный анализ индукции ИФН-а в сыворотке крови и легочной ткани мышей препаратом Амиксин в дозе 400 мг/кг показал, что его продукция в легочной ткани значительно (в 2,8 раза) выше, чем в сыворотке крови. Однако пик продукции ИФНа в сыворотке крови определялся через 24 ч после однократного введения препарата, максимальная же продукция ИФН-а в легочной ткани мышей тестировалась только через 48 ч. Иными словами, продукция ИФНа под действием Амиксина определялась сначала в сыворотке крови (через 24 ч после введения), а затем — в легочной ткани (через 48 ч), но в легочной ткани ИФН-а было больше (рис. 5).
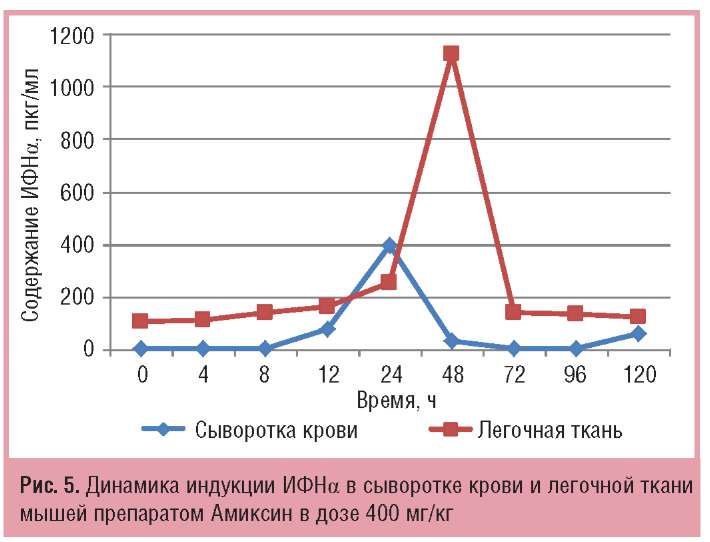
Максимальный уровень продукции ИФН-λ2/3 в легочной ткани мышей под действием Амиксина в дозе 400 мг/кг достигался через 12 ч после введения препарата и составлял 1547,5+3,7 пкг/мг, а пик его продукции в сыворотке крови тестировался через 24 ч после введения препарата и составлял 2231+93 пкг/мл, что превышало содержание ИФНλ2/3 в легочной ткани на пике его продукции в 1,4 раза. Иными словами, максимальный синтез ИФН-λ2/3 в легочной ткани достигался быстрее, чем в сыворотке крови мышей, но его количество было меньше (рис. 6).
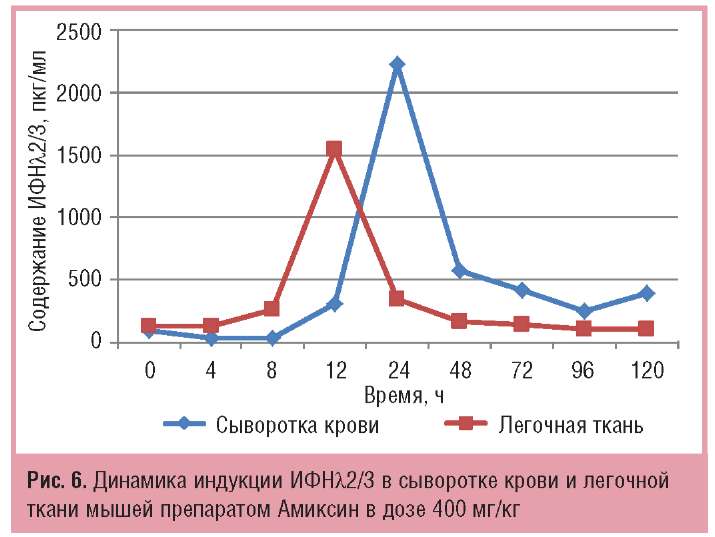
Таким образом, индукция и продукция ИФНа в сыворотке крови опережает таковую в легочной ткани на 24 ч, но на пике продукции количество ИФН-а в сыворотке крови значительно уступает таковому в легочной ткани (в 2,8 раза), а индукция и продукция ИФНλ2/3 в легочной ткани опережает таковую в сыворотке крови мышей на 12 ч, но на пике продукции количественно тестируется на более низком уровне, чем в сыворотке крови мышей (в 1,4 раза; см. рис. 6).
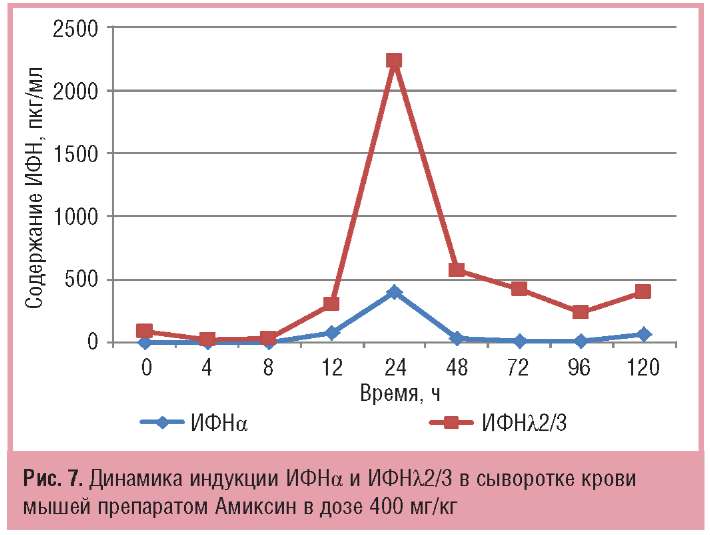
Сравнительный анализ результатов индукции ИФНа и -λ2/3 в сыворотке крови мышей под действием однократной дозы Амиксина 400 мг/кг показал, что срок достижения их максимальной продукции совпадает — через 24 ч после введения препарата. Однако количество в крови ИФНλ2/3 на пике его продукции в 5,6 раза выше, чем ИФНа (соответственно 2231+93 и 398+12 пкг/мл; рис. 7).
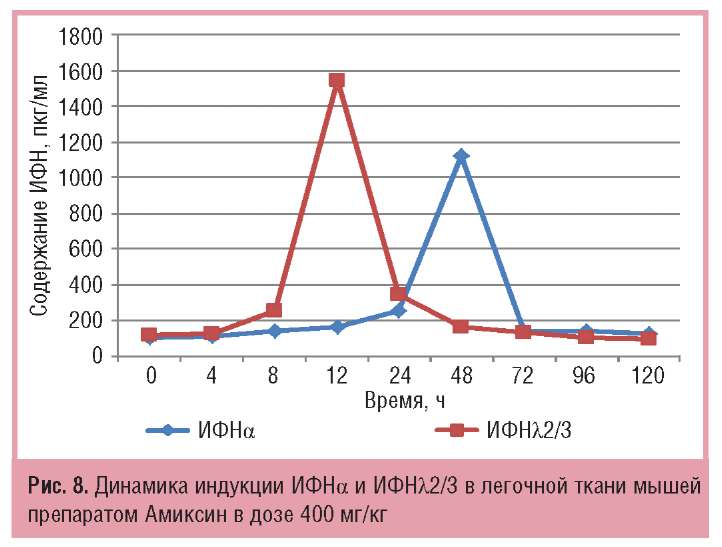
В легочной ткани мышей максимальный синтез ИФНλ2/3 под действием Амиксина в той же дозе опережает синтез ИФНа на 36 ч и превосходит его количественно в 1,4 раза (соответственно 1547+3 и 1125,00+14,43 пкг/мг; рис. 8).
Итак, исходя из суммарных данных исследования и их сравнительного анализа, препарат Амиксин в дозе 40 мг/кг (эквивалентной суточной дозе для человека) вызывает пролонгированную системную индукцию и продукцию ИФНа, -β, -γ и- λ в сыворотке крови, максимальный уровень которых определяется уже через 24 ч после введения препарата, а фоновые значения сохраняются до 120 ч (5 сут) включительно. Одновременно Амиксин в дозе 40 мг/кг индуцирует пролонгированную локальную продукцию ИФНλ, и α в легочной ткани экспериментальных животных, максимальный уровень которой тестируется соответственно через 24 и 48 ч после введения препарата и также сохраняется на терапевтических значениях до 96 и 120 ч включительно.
Динамика аккумуляции ИФНа, β, -γ и - λ в крови при использовании препарата Амиксин в дозе 400 мг/кг (эквивалентной курсовой для людей) аналогична таковой при дозе 40 мг/кг, но количества ИФНа и -Я, синтезированных на пике продукции, увеличиваются в среднем в 2 раза, а ИФНр и -γ — в 10 раз и сохраняются на уровне терапевтических значений в течение 120 ч. Максимальная локальная продукция ИФНа в легочной ткани мышей под действием Амиксина в дозе 400 мг/кг определяется через 48 ч, а синтез максимальных количеств ИФНλ2/3 увеличивается и ускоряется на 36 ч, опережая максимальную продукцию ИФНа в легочной ткани при дозе препарата 400 мг/кг. В последующие сроки исследования до 120 ч включительно в легочной ткани экспериментальных животных определяется локальная продукция терапевтического количества как ИФНа, так и ИФНλ.
Таким образом, однократный прием препарата Амиксин в дозе, эквивалентной суточной дозе для человека, в течение первых 24 ч индуцирует ИФН 1-го, 2-го и 3-го типов в крови и легочной ткани мышей. Повторные приемы Амиксина в указанной дозе (суммарно соответствующей курсовой дозе для человека) с интервалом в 48 ч позволяют избежать характерного для индукторов ИФН феномена гипореактивности и обеспечивают постоянную максимальную выработку всех типов ИФН в крови и дополнительно локальную — в легочной ткани. Следует отметить также, что, хотя по результатам исследований максимальная продукция ИФНа и -Я2/3 в сыворотке крови мышей под действием Aмиксинa в дозах 40 и 400 мг/кг тестируется одновременно — через 24 ч, количество ИФНλ2/3 выше, чем ИФНа, соответственно в 7,6 и 5,6 раза. В легочной ткани мышей также всегда тестируется большее содержание ИФНλ2/3, сроки достижения которого опережают таковые для ИФНа соответственно на 24 и 36 ч. Такая последовательность системной и локальной продукции ИФНа и - λ в сыворотке крови и легочной ткани указывает на взаимно индуцирующий характер их взаимодействия, когда предварительно синтезированный в большем количестве ИФНλ2/3 способствует и (или) стимулирует индукцию и продукцию ИФНа и наоборот. Известно, что ИФНа и - λ имеют общий механизм регуляции продукции и обладают взаимно индуцирующим действием [20—22]. Многочисленными исследованиями последних лет установлено также, что разные штаммы вируса гриппа А, в том числе высокопатогенный штамм H1N1 (2009), обладают повышенной чувствительностью к ИФН-α и - λ [11, 15, 23, 29], преобладающая продукция которых определяется и в проведенных нами исследованиях. Индуцированные препаратом Амиксин плюрипотентные молекулы ИФН 1-го, 2-го и 3-го типов могут оказывать как системное действие на организм в целом, так и, что особенно важно, адресное противовирусное действие на орган-мишень не только при гриппе, но и при других острых респираторных вирусных инфекциях (ОРВИ) и выполняют ключевую роль в повышении барьерной защитной функции легочной ткани [12, 24—28, 30]. Согласно последним научным публикациям, при инфицировании вирусом гриппа А количество вирусиндуцированного ИФНА, в легочной ткани мышей также больше, чем ИФНа [11, 12, 23], что указывает на его доминантную, защитную роль в очагах поражения гриппозной инфекцией.
Корреляция результатов действия разных доз препарата Амиксин в условиях отсутствия инфицирования с аналогичными данными под действием вируса гриппа А связана, вероятно, со сходством механизмов продукции ИФНа и -Я, индуцированных вирусом и препаратом Амиксин, что научно обосновывает профилактический и лечебный эффекты препарата, направленные на повышение противовирусной защиты «входных ворот» инфекции, органов-мишеней и организма в целом как при гриппозной, так и при других ОРВИ. Не исключено также, что подобный механизм преобладания индукции Амиксином локальной продукции ИФНа и -Я, особенно ИФНλ, во «входных воротах» инфекции и органах-мишенях может быть актуален при вирусных инфекциях кишечного и урогенитального трактов. Ведущая защитная роль локальной продукции ИФНλ описана, в частности, при ротавирусной и генитальной герпетической инфекциях [10, 11, 14]. Ранее при изучении кинетики синтеза под действием Амиксина ИФН в органах и тканях также установлена опережающая во времени и количественно его продукция в такой последовательности: кишечник—печень—легкие—кровь [3, 17].
В настоящей работе впервые в строго контролируемых лабораторных условиях, соответствующих принципам доказательной медицины, идентифицированы все 3 известных типа ИФН, которые индуцируются препаратом Амиксин в крови и легочной ткани; подробно изучены динамика, последовательность и профиль их системной и локальной продукции. Согласно результатам исследования, препарат Амиксин является быстродействующим (в первые 24 ч) противовирусным средством эндогенной пролонгированной стимуляции выработки ИФН 1—3-го типов в крови и ИФН 1-го и 3-го типов — в легочной ткани. В совокупности они активируют собственные защитные силы легких и организма в целом, что свидетельствует о возможности профилактического применения препарата для предотвращения повторных ОРВИ. Преобладание аккумуляции ИФНа и - λ в крови и, что особенно важно, — ИФНА, в легких, указывающее на его доминирующую роль в защите легочной ткани — «входных ворот» респираторной инфекции разного генеза — свидетельствует о том, что препарат Амиксин может и должен использоваться с лечебной целью как на ранних, так и на поздних стадиях развития ОРВИ для противовирусной защиты 1-й линии от воздействия возбудителя на орган-мишень. Многократное профилактическое или лечебное применение препарата Амиксин с минимальным интервалом в 48 ч позволит ускорить и увеличить продукцию ИФН и избежать развития феномена гипореактивности.
Таким образом, сегодня препарат Амиксин является единственным из зарегистрированных в РФ индукторов ИФН, действие которого на выработку всех 3 типов ИФН в крови и 1-го и 3-го типов — в легочной ткани достоверно доказано.
Индукция препаратом Амиксин эндогенных ИФН 1—3-го типов в крови и легочной ткани активирует врожденный и адаптивный иммунный ответ, сохраняет иммунный гомеостаз организма и предотвращает и (или) прерывает синтез вирусных нуклеиновых кислот и вирусспецифических белков, не вызывая характерных для экзогенных препаратов ИФН побочных эффектов.
Литература
1. Mayer G., Kruger R. Tilorone hydrochloride: mode de action // Science. -1970; 169: 1214-5.2. Чижов Н.П. Смольская Т.Т., Байченко П.И. и др. Клинические исследования переносимости и интерферон индуцирующей активности амиксина // Вопр. вирусологии. - 1990; 5: 411—1.
3. Тазулахова Э.Б. Закономерности индукции и продукции а-, у- интерферона. Автореф. дис. ... д-ра биол. наук. М., 1986.
4. Интерферон-2011. Сб. научных статей / М., 2012.
5. Shapira S., Hacohen N. Systems biology approaches to dissect mammalian innate immunity // Curr. Opin. Immunol. - 2011; 23: 2373-7.
6. Hertzog P. Type 1 interferons as primers, activators and inhibitors of innate and adaptive immune responses // Immunol. Cell. Biol. - 2012; 90: 471-3.
7. Hervas-Stubs S., Perez-Garcia J. et al. Direct effects of type 1 interferons on cеlls of immune system // Clin. Cancer Res. - 2011; 17: 2619-27.
8. Choubey D., Moudgi K. Interferons in autoimmune and inflammatory diseases: regulation and roles // J. Interferon & Cytokine Res. - 2011; 12: 857-65.
9. Hillyer Ph., Mane V. et al. Expression profiles of human interferon-alpha and interferon-lambda subtypes are ligand- and cell-dependent // Immunol. Cell Biol. - 2012; 90: 774-83.
10. Mordstein M., Neugebauer E. et al. Lambda Interferon reders epithelial cells of respiratory and gastrointestinal tracts resistant to viral infections // J. Virol. -2010; 11: 5670-77.
11. Durbin R., Kotenko S., Durbin J. Interferon induction and function at the mucosal surface // Immunol. Rev. - 2013; 255: 25-39.
12. Jewell N., Cline T. et al. Lambda Interferon is the predominant interferon induced by Influenza A virus infection in vivo // J. Virol. - 2010; 84: 11515-22.
13. Donnelly R., Kotenko S. Interferon-Lambda: A new addition to an old Family // J. Interferon & Cytokine Res. - 2010; 8: 555-64.
14. Kotenko S. IFN-Xs // Curr. Opin. Immunol. - 2011; 23: 1-8.
15. Levy D., Marie I. Induction of type 1 and 3 interferon in response to viral infection // Curr. Opin. Virol. - 2011; 1: 476-86.
16. Wang B., Fish E. The yen and yang of viruses and interferons // Trend Immunol. - 2012; 33: 190-7.
17. Григорян С.С., Ершов Ф.И., Поверенный А.М. и др. Особенности продукции интерферона при энтеральном введении индукторов интерферона // Вопр. вирусол. - 1988; 1: 67-70.
18. Григорян С.С., Иванова А.М., Ершов Ф.И. Противовирусная активность амиксина и его влияние на интерфероновый статус // Вопр. вирусол. - 1990; 2: 138-Ю.
19. Миронов А.Н., Бунатян Н.Д. и др. Руководство по проведению доклинических исследований лекарственных средств / М.: Гриф и К, 2012.
20. Iversen M., Paludan S. Mechanisms of type 3 interferon expression // J. Interferon & Cytokine Res. - 2010; 8: 573-8.
21. Onoguchi K., Yoneyama M. et al. Viral infection activate type 1 and 3 interferon gene through a common mechanism // J. Biol. Chem. - 2007; 282: 7576-81.
22. Ank N., West H. et al. X Interferon (IFN-A), a type 3 IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo // J. Virol. - 2006; 80: 4501-9.
23. Hermant P., Michiels Th. Interferon-X in the context of viral infections; production, response and therapeutic implication // J. Innate Immune. - 2014; 6: 563-74.
24. Okabayashi T., Kojima T. Masaki T. et al. Type 3 interferon, not type 1, is predominant interferon induced by respiratory viruses in nasal epithelial cells // Virus Res. - 2011; 1-2: 360-6.
25. Mordsstein M., Neugebauer E., Ditt V. et al. Lambda interferon renders epithelial cells of respiratory and gastrointestinal tracts resistant to viral infections // J. Virol. - 2010; 84 (11): 5670-7.
26. Svetlikova D., Kabat P. et al. Influenza A virus replication is inhibited in IFN -Л2 and IFN-A3 transfected or stimulated cells // Antiviral. Res. - 2010; 88 (3): 329-3.
27. Khaitov M., Laza-Stanca V., Edwards M. et al. Respiratory virus induction of alpha-beta- and lambda interferons in bronchial epithelial cells and peripheral blood mononuclear cells // Allergy. - 2009; 63 (3): 375-86.
28. Wang J., Oberley-Deegan R., Wang Sh. et al. Differentiated human alveolar type 2 cells secrete antiviral IL-29 (IFN-A1) in response to influenza A infection // J. Immunol. - 2012; 182: 1296-304.
29. Osterlund P., Pirhonen J., Ikonen N. et al. Pandemic H1N1 Influenza A virus highly sensitive to the antiviral action of interferons // J. Virol. - 2010; 84: 1414-22.
30. Ank N., Iversen M. et al. An important role for type 3 interferon in TLR-induced antiviral activity // J. Immunol. - 2008; 180: 2474-85.
Комментарии
ПРАКТИКА ПЕДИАТРА



