Ультраструктурные нарушения отростков астроцитов в префронтальной и зрительной коре при шизофрении
СтатьиВихрева О.В.1, Рахманова В.И.1, Клинцова А.Ю.2,3, Гринау У.Т.2, Уранова Н.А.1 1 ФГБНУ НЦПЗ, Москва, Россия; 2 Университет штата Иллиноис, США; 3 Университет штата Делавер, США
Цель: провести морфометрическое электронномикроскопическое исследование отростков астроцитов (ОА) в префронтальной коре (ПФК) и зрительной коре (ЗК), включая ОА, прилежащие к синапсам на шипиках дендритов, в ПФК и ЗК при шизофрении и в контроле.
Материал и методы: исследованы ПФК (поле 10 по Бродману) и ЗК (поле 17 по Бродману) в 20 аутопсийных случаях шизофрении и 16 нормальных контрольных случаях. Определяли объемную фракцию (Vv) ОА и содержащихся в них митохондрий в слоях I и II в группах шизофрении и контрольной.
Результаты: в ПФК в слое I показано снижение Vv ОА (-31%), содержащихся в них митохондрий (-48%, p < 0,05) и Vv ОА, прилежащих к аксо-шипиковым синапсам (-44%, p < 0,05) при шизофрении по сравнению с контролем. В ЗК в слое I значимых изменений не найдено. В слое II только в ЗК показано снижение Vv ОА (-38%, p < 0,01).
Заключение: регрессивное снижение объемной фракции ОА при шизофрении регионально специфично, может быть связано со снижением численной плотности аксо-шипиковых синапсов и нарушением функциональной активности в ПФК. Эти изменения могут быть результатом дизонтогенеза при шизофрении.
Ключевые слова: отростки астроцитов; аутопсия; ультраструктура; морфометрия; префронтальная кора; зрительная кора; шизофрения
Для цитирования: Вихрева О.В., Рахманова В.И., Клинцова А.Ю., Гринау У.Т., Уранова Н.А. Ультраструктурные нарушения отростков астроцитов в префронтальной и зрительной коре при шизофрении // Психиатрия. 2018. № 79 (3). С.
Конфликт интересов отсутствует.
Ultrastructural abnormalities of astrocytic cell processes in prefrontal and visual cortex in schizophrenia
Vikhreva O.V.1, Rakhmanova V.I.1, Klintsova A.Ju.2,3, Greenough W.T.2, Uranova N.A.1
1 Federal State Budgetary Scientific Institution Mental Health Research Centre, Moscow, Russia; 2 University of Illinois at Urbana-Champaign, USA; 3 University of Delaware, USA
Aim: to perform electron microscopic morphometric study of astrocytic processes (AP), including those directly adjacenttoaxo-spinous synapses, in the prefrontal cortex (PFC) and visual cortex (VC) in postmortem brains of schizophrenia patients and normal controls.
Material and methods: PFC and VC (Brodmann areas 10 and 17) obtained from autopsies of 20 cases of schizophrenia and 16 normal controls. Volume fraction (Vv) of APand Vv of mitochondria inside these processes, including those adjacentto axo-spinous synapses, were estimated layers I and II of the PFC and VC in the schizophrenia and control groups.
Results: in layer I of the PFC, a significant decrease in Vv of AP (-31%, p < 0,05), of AP mitochondria (-48%, p < 0,05) and Vv of AP adjacentto axo-spinous synapses (-44%, p < 0,05) were found in schizophrenia as compared to controls. No significant changes in any of those parameters were detected in the VC of the schizophrenia groupcompared with control group. In layer II a significantly lower Vv of AP were found only in the VC in schizophrenia (-38%, p < 0,01).
Conclusion: regressive decrease in the Vv of AP in schizophrenia is region-specific and might be associated with the decrease in the density and the dysfunction of axo-spinous synapses in the PFC. Thechange smight be of development origin.
Keywords: astrocytic cell processes; post-mortem; ultrastructure; morphometry; prefrontal cortex; visual cortex; schizophrenia For citation: Vikhreva O.V., Rakhmanova V.I., Klintsova A.Ju., Greenough W.I., Uranova N.A. Ultrastructural abnormalities of astrocytic cell processes in prefrontal and visual cortex in schizophrenia // Psychiatry. 2018. № 79 (3). P.
There is no conflict of interest.
Введение
Префронтальная кора (ПФК) является одним из главных объектов изучения мозга при шизофрении. С нарушением ее функций и особенно синаптических контактов связывают расстройства мышления, мотиваций и эмоциональности, позитивные и негативные симптомы шизофрении [1]. По данным нейровизуализационных и аутопсийных исследований структурные изменения в ПФК при шизофрении включают снижение объема серого вещества [2], отсутствие реактивного глиоза [3, 4], дефицит олигодендроглии [5, 6], атрофию дендритов нейронов, потерю дендритных шипиков при отсутствии изменений в числе нейронов [7-9]. В ПФК при шизофрении показаны снижение экспрессии генов, вовлеченных в синаптическую передачу, дисфункция (включая дефицит рабочей памяти), связанная с нарушением нейронных микросетей в ПФК и связей ПФК с другими структурами мозга [10]. Эти изменения дали основание предполагать нарушение организации синаптических связей в виде изменения их численной плотности на шипиках и стволах дендритов в ПФК при шизофрении. Ультраструктурные морфометрические исследования аутопсийного мозга показали снижение размеров дендритных шипиков [11, 12], числа митохондрий в синапсах [13-16]. Ранее нами были обнаружены изменения численной плотности синапсов на шипиках дендритов в ПФК при шизофрении [17]. Причина описанных нарушений остается неясной. Предполагается, что они могут быть следствием дефекта нейронной миграции, повреждения миелинизации и запрограммированной элиминации синапсов во время развития мозга.
Интерес к астроцитам при шизофрении обусловлен тем, что эти клетки структурно и функционально тесно связаны с нейронами и участвуют в механизмах синаптической передачи и пластичности, в энергетическом и глутаматном метаболизме нейронов, в ионном гомеостазе [18]. Дисфункция астроцитов и нарушения их энергетического, глутаматного и кинуренинового метаболизма в мозге при шизофрении показаны в биохимических и молекулярно-генетических исследованиях [19-21]. При шизофрении в ПФК обнаружены как снижение содержания и экспрессии основного маркера астроцитов — глиального фибриллярного кислого белка [22, 23], так и повышенная экспрессия этого белка [24] по сравнению с контролем. Оказалось, что найденный в некоторых исследованиях глиоз при шизофрении сочетается с нейровоспалением [25]. Ультраструктурные дистрофические нарушения астроцитов, связанные со снижением численной плотности и объемной фракции митохондрий и накоплением липофусцина, показаны при шизофрении в гиппокампе [26].
Астроциты участвуют в синаптогенезе, могут элиминировать дендритные шипики, регулировать эффективность синаптической передачи и нейрональную пластичность [18]. Синаптические контакты метаболически и функционально связаны с астроцитами с помощью расположенных на них рецепторов к нейротрансмиттерам, отростки астроцитов (ОА) тесно прилежат к пре- и постсинаптическим компонентам синапсов [27]. Синаптическая структурная пластичность происходит параллельно с изменениями в ОА при длительной пост-тетанической потенциации [18], пребывании животных в обогащенной среде [28, 29]. Ранее нами было показано значимое снижение численной плотности синапсов на шипиках дендритов в слое I, повышение в слое II ПФК и отсутствие изменений в зрительной коре (ЗК) при шизофрении [17] и дистрофические изменения астроцитов в верхних слоях ПФК при шизофрении [30]. Rajkowska et al. [3] показали снижение площади, занимаемой астроцитами и их отростками в слое V ПФК при шизофрении. Такого рода нарушения могут быть обусловлены регрессивными изменениями и/или гипотрофией связанных с синапсами ОА при шизофрении, что может проявляться в их ультраструктурных нарушениях, включая отростки, непосредственно связанные с синапсами.
Целью настоящей работы было провести морфометрическое электронномикроскопическое исследование ОА в нейропиле и содержащихся в них митохондрий, включая ОА, прилежащие к синапсам на шипиках дендритов, в ассоциативной ПФК и проекционной ЗК при шизофрении и в контроле.
Материал и методы
Исследование проведено на коллекции мозговых структур аутопсийного мозга лаборатории клинической нейроморфологии ФГБНУ «Научный центр психического здоровья». Диагностику шизофрении при жизни проводили по МКБ-10. Исследованы ПФК (поле 10 по Бродману) в 20 случаях шизофрении и 16 нормальных контрольных случаях и ЗК (поле 17 по Бродману) в 18 случаях шизофрении и 16 контрольных. Группа шизофрении включала 20 случаев параноидной шизофрении: 11 случаев c приступообразно-прогредиентным течением (F20.01) и девять случаев c непрерывным типом течения (рубрика F20.00). Контрольная группа включала 16 случаев без психической патологии. Характеристика группы шизофрении и контрольной группы дана в табл. 1. Причины смерти были сходными в обеих группах: инфаркт миокарда, тромбоэмболия легочной артерии, пневмония, ишемическая болезнь сердца, острая сердечно-сосудистая недостаточность. Терапию нейролептиками оценивали в виде хлорпромазинового эквивалента [31, 32]. Предварительное нейрогистологическое исследование всех образцов мозговой ткани показало отсутствие признаков болезни Альцгеймера, сосудистых заболеваний, паркинсонизма.
Таблица 1
Демографическая характеристика групп
|
Контрольная группа (n = 16) |
Группа шизофрении |
|
|
Возраст, лет |
49,18 ± 13,0 |
57,65 ± 16,0 |
|
Постмортальный интервал (часы) |
5,4 ± 1,0 |
5,7 ± 2,32 |
|
М/ж |
15/1 |
5/15 |
Была изучена ткань ПФК (поле 10) и зрительной коры (поле 17). Для каждого случая кусочки ткани брали из левого полушария, фиксировали погружением в смесь растворов 2,5% глутаральдегида и 4% параформальдегида на 0,1М фосфатном буфере в течение 1 нед. Из фиксированной ткани вырезали кусочки толщиной 0,5 мм через все слои коры перпендикулярно поверхности, промывали в фосфатном буфере, дофиксировали осмием 1ч, дегидратировали и заливали в эпоксидную смолу Аралдит. Полутонкие срезы (1 мкм), окрашенные метиленовым синим, использовали для ориентации в слоях коры. Слои ПФК и ЗК мозга человека идентифицировали по известным цитоархитектоническим критериям. Использовали три случайно выбранных блока ткани на случай из каждой исследованной структуры мозга. Ультратонкие срезы собирали на медные бленды, покрытые подложкой из формвара и стабилизированные углеродным напылением. Ультратонкие срезы окрашивали водным раствором уранилацетата и раствором цитрата свинца и просматривали с помощью электронного микроскопа Philips EM-420 (Голландия).
Морфометрическое исследование проводили слепым методом. Определяли объемную фракцию (Vv) ОА в нейропиле и содержащихся в них митохондрий, включая отростки, прилежащие к аксо-шипиковым синапсам, в слоях I и II при шизофрении и в контроле без психической патологии. Фотографии нейропиля каждого из двух слоев ПФК и ЗК получали при увеличении 6,500х. Морфометрию проводили на приборе «Minolta» на негативах при конечном увеличении 50,800х и с помощью наложенных на них тестовых сеток [38]. Площадь каждой микрофотографии была 126 мкм2, общая исследованная площадь на случай составляла в среднем 3024 мкм2. Отростки астроцитов в нейропиле легко идентифицировали по их электронно-прозрачной цитоплазме, неправильной форме, отсутствию органелл, за исключением митохондрий, и синаптических мембранных специализаций и тесному соприкосновению с синапсами [33].
Объемную фракцию (Vv) астроцитарных отростков, в том числе прилежащих к синаптическим контактам, оценивали с помощью стереологического метода [34]. Для этого прозрачную квадратную тестовую сетку накладывали на негатив изображения нейропиля и подсчитывали число точек на тестовой сетке, приходящихся на ОА. Vv астроцитарных отростков определяли как число этих точек, деленное на общее число точек тестовой сетки, приходящихся на всю площадь негатива и выраженное в процентах. Vv митохондрий в астроцитарных отростках определяли как процент точек, приходящихся на митохондрии, ко всем точкам на астроцитарном отростке. Определяли средние значения параметров для каждого негатива и затем для каждого случая с последующей оценкой средних групповых значений.
Статистический анализ проводили с помощью пакета STATISTICA 6. Нормальность распределения оцениваемых параметров в группах тестировали по критерию Колмогорова-Смирнова. Для выявления возможного влияния возраста, постмортального интервала и хлор-промазинового эквивалента, длительности болезни, возраста начала болезни применяли корреляционный анализ Пирсона. Влияние пола оценивали с помощью двухфакторного дисперсионного анализа. Для групповых сравнений использовали однофакторный дисперсионный анализ и трехфакторный дисперсионный анализ с диагнозом, возрастом и постмортальным интервалом с последующим posthoc тестом (Duncan test, p < 0,05).
Результаты и обсуждение
Электронномикроскопическое исследование показало хорошую сохранность ткани ПФК и ЗК в норме и при шизофрении (рис. 1).
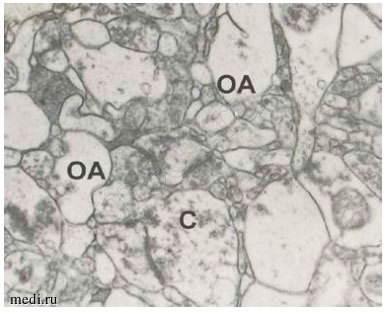 Рис. 1. Отростки астроцитов (ОА,*) и аксо-шипиковые синапсы (С) в нейропиле слоя I префронтальной коры в контроле (А) и при шизофрении (Б). Масштаб — 0,5 мкм
Рис. 1. Отростки астроцитов (ОА,*) и аксо-шипиковые синапсы (С) в нейропиле слоя I префронтальной коры в контроле (А) и при шизофрении (Б). Масштаб — 0,5 мкм
Префронтальная кора. Статистический однофакторный дисперсионный анализ показал значимый эффект диагноза на Vv ОА (-31%): F (1,34) = 5.01, p = 0.03; Vv митохондрий в ОА (-48%): F (1,34) = 4,61, p = 0,04) и Vv ОА, прилежащих к аксо-шипиковым синапсам (-44%): F (1,34) = 5.07, p = 0,03) в слое I и статистически незначимое снижение этих параметров в слое II (табл. 2, рис. 2). Трехфакторный дисперсионный анализ подтвердил главный эффект диагноза на следующие параметры в слое I: Vv отростков астроцитов F (1, 28) = 5,81, p = 0,02, Vv митохондрий в ОА F (1,28) = 6,1, p = 0,02 и Vv ОА, прилежащих к аксо-шипиковым синапсам F (1,28) = 9,25, p = 0,005. Posthoc тест выявил значимые групповые различия по этим параметрам. Не найдено влияния возраста, постмортального интервала, пола, хлорпромазинового эквивалента, длительности болезни, возраста начала болезни на измеренные параметры.
Таблица 2
Средние и ст. ошибка среднего объемной фракции (Vv) ОА и содержащихся в них митохондрий в слоях I и II ПФК и ЗК в контрольной группе и в группе шизофрении
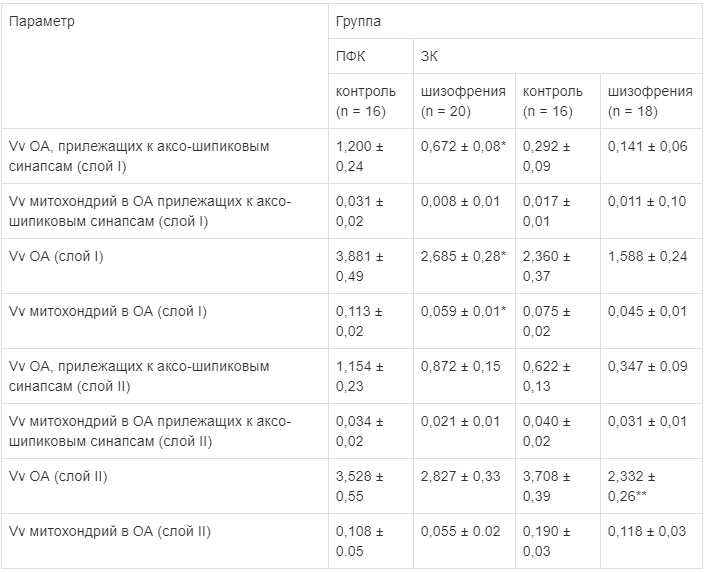
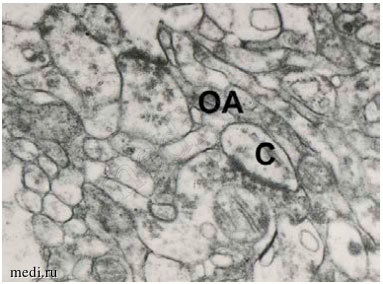 Рис. 2. Слой I префронтальной коры и зрительной коры. Объемная фракция отростков астроцитов (ОА), прилежащих к синапсам (А), отростков астроцитов (Б) и митохондрий в отростках астроцитов (В) в контроле и при шизофрении
Рис. 2. Слой I префронтальной коры и зрительной коры. Объемная фракция отростков астроцитов (ОА), прилежащих к синапсам (А), отростков астроцитов (Б) и митохондрий в отростках астроцитов (В) в контроле и при шизофрении
Зрительная кора. Однофакторный дисперсионный анализ показал значимый эффект диагноза на Vv ОА в слое II (-37%): F (1.32) = 8.89, p < 0,01 и отсутствие значимых различий в других параметрах и в слое I (табл. 2, рис. 3). Трехфакторный дисперсионный анализ подтвердил главный эффект диагноза на Vv ОА в слое II (p < 0,01). Не найдено влияния возраста, постмортального интервала, пола, хлорпромазинового эквивалента, длительности болезни, возраста начала болезни на измеренные параметры.
Рис. 3. Объемная фракция отростков астроцитов (ОА) в слое II префронтальной и зрительной коры в контроле и при шизофрении
Таким образом, в проведенном исследовании было установлено значимое снижение объемной фракции ОА в нейропиле, содержащихся в них митохондрий и ОА, прилежащих к аксо-шипиковым синапсам, в слое I ПФК. При этом в ЗК в слое I значимых изменений не найдено. В слое II только в ЗК показано снижение Vv ОА. Статистический анализ показал, что эти изменения не связаны с возрастом и постмортальным интервалом. Полученные данные согласуются с результатами аутопсийного исследования Rajkowska и соавт. [3], обнаруживших снижение площади, занимаемой астроцитами и их отростками, в слое V ПФК при шизофрении.
Важным вопросом является влияние нейролептической терапии на астроциты. Konopaske и соавт. [35] вывили снижение числа S100B-иммунореактивных астроцитов в париетальной коре приматов под воздействием галоперидола и оланзапина. Однако неизвестно, снижались ли при этом размеры астроцитов. В нашем исследовании не было найдено корреляции астроцитарных параметров с хлорпромазиновым эквивалентом нейролептичекой терапии. Ранее нами было показано, что 28-недельное лечение больных шизофренией оланзапином вызывало в органотипической культуре ткани мозга человека гипертрофию астроцитов и повышение в них числа митохондрий [36]. Эти данные дают основание полагать, что обнаруженными в настоящем исследовании нарушения не связаны с действием антипсихотической терапии. К сожалению, мы не смогли оценить влияние пола на исследованные параметры, поскольку нам не удалось адекватно подобрать группу по полу.
Реактивность астроцитов на структурном уровне проявляется в изменении их размеров, отражающем гипертрофию или атрофию этих клеток и их отростков. Наше исследование подтвердило высказанное нами предположение о регрессивных изменениях ОА при шизофрении и их возможной связи с изменения синапсов, поскольку ранее было показано снижение численной плотности синапсов на шипиках дендритов слое I ПФК [17] и размеров аксошипиковых синапсов в поле 10 ПФК [12] при шизофрении по сравнению с контролем, указывающие на снижение эффективности синаптической передачи [16]. Эти результаты согласуются со снижением числа шипиков и атрофией дендритного дерева нейронов, в том числе в поле X ПФК при шизофрении на препаратах, окрашенных по методу Гольджи [8].
Важно отметить, что снижение объемной фракции ОА сопровождалось снижением содержащихся в них митохондрий в ПФК при шизофрении по сравнению с контролем. Нарушение регуляции биоэнергетического взаимодействия астроцитов и синапсов является одним из наиболее важных механизмов патогенеза шизофрении [37]. В наших предыдущих исследованиях выявлен дефицит митохондрий в астроцитах в гиппокампе [26], стриатуме [15] при шизофрении, что было подтверждено работами других авторов [14, 16]. В гиппокампе при шизофрении показано снижение объемной фракции и числа митохондрий в астроцитах, негативно коррелирующее с длительностью болезни [26].
Дефицит митохондрий в астроцитарных отростках, выявленный в настоящем исследовании, может быть связан с нарушением митохондриального дыхательного окислительного фосфорилирования при шизофрении [38], специфическими для шизофрении нарушениями митохондриального комплекса I в ПФК аутопсийного мозга [39] и выявленной при шизофрении дисфункцией митохондрий и подвержености оксидативному стрессу [40].
Проведенное исследование также показало, что изменения ОА регионально специфичны, поскольку они найдены в слое I ПФК и в слое II ЗК при шизофрении. В связи с этим интересно отметить, что с помощью прижизненных методов исследования, таких как позитронная эмиссионная томография, функциональная ядерно-магнитная резонансная томография установлены функциональные нарушения ассоциативных связей ПФК с теменной корой, передней лимбической корой, таламусом и гиппокампом [1]. ПФК играет центральную роль в социальных контактах, будучи связанной с височной и теменной корой и подкорковыми структурами, и изменения этих контактов у больных шизофренией связаны с нарушением функциональных взаимосвязей этих корковых областей. Снижение объемной фракции ОА в ассоциативном слое II ЗК при шизофрении согласуется с дефицитом сенсорного зрительного восприятия объектов, нарушением ассоциативных зрительных функций, связанных с нарушением внутренних связей в зрительной системе [41] и снижением функционирования межполушарных связей в зрительной системе при шизофрении [42].
Возникает вопрос, с какими синаптическими нейротрансмиттерами могут быть связаны обнаруженные изменения астроцитарных отростков? Астроциты активно участвуют в энергетическом и глутаматном метаболизме. В настоящее время астроциты рассматриваются как неотъемлемые компоненты глутаматергических синапсов, состоящие из пре- и постсинапса и астроцитарного отростка, прилежащего к синапсу. С помощью методов иммуноцитохимии установлено, что наиболее многочисленные аксошипиковые синапсы в верхних слоях ПФК образованы внутренними коллатералями пирамидных нейронов слоя III на других пирамидных клетках, возбуждающими ассоциативными волокнами, глутаматными аксонными терминалями [43]. Электронномикроскопическое иммуноцитохимическое исследование аутопсийной ПФК обнаружило преимущественное расположение транспортеров возбуждающих аминокислот (глутамата) на мембранах астроцитарных отростков и в непосредственной близости от синапсов [44]. При шизофрении в ПФК показано связанное с когнитивными нарушениями [46] повышение экспрессии одного из генов глутаматных NMDA-рецепторов, экспрессии mGIuR1a- и mGluR2/3-рецепторов, плотности AMPA-глутаматных рецепторов, которые локализуются в возбуждающих синапсах [45], и концентрации глутамата, а также снижение уровня связывания 3Н-каината во всех слоях ПФК, включая слои I—II [47]. В аутопсийном мозге больных шизофренией выявлены нарушения экспрессии и/или активности астроцитарных глутаминсинтетазы и переносчиков глутамата (преимущественно в ПФК и передней поясной извилине) [23]. Эти данные позволяют предполагать, что изменения ОА в ПФК могут быть связаны прежде всего с нарушениями в глутаматергических синапсах при шизофрении. К другим наиболее значимым нейротрансмиттерным системам, участвующим в изменении численной плотности синаптических связей в ПФК при шизофрении, могут быть ГАМКергические синапсы, дофаминергические синапсы, норадренергические синаптические контакты.
Снижение объемной фракции ОА (-30-40%) наряду с установленным нами ранее снижением числа и размеров самых многочисленных аксо-шипиковых синаптических контактов в ПФК при шизофрении вносит свой вклад в снижение объема нейропиля и в целом объема серого вещества ПФК при шизофрении [2]. Однако причина этого снижения остается неизвестной. Goudriaan и соавт. [48] в генетическом ассоциативном исследовании показали, что повышенный риск развития шизофрении связан с генами астроцитов и олигодендроцитов. Молекулярно-генетические исследования показали нарушения экспрессии астроцитарных генов в ПФК при шизофрении, влияющих на реактивность этих клеток, особенно генов ростовых и нейротрофических факторов и их рецепторов. В работе Hagihara и соавт. [49] было установлено, что паттерн изменения экспрессии генов в ПФК при шизофрении напоминает паттерн ювенильной ПФК. В связи с этим интересно отметить, что высокая экспрессия ГФКБ характерна для зрелых астроцитов, а в ПФК при шизофрении показано снижение экспрессии ГФКБ, что, возможно, связано с их незрелостью. Markham и соавт. [50] показали, что созревание ПФК в пубертатный период связано с полом и нарушается пренатальным стрессом. ПФК развивается сравнительно поздно, и в пубертатный период глиальные клетки играют большую роль в созревании мозга, особенно в синаптогенезе и пластичности, а также во взаимоотношении синапсов с астроцитами. Возможно, гормональная перестройка в этот период может вызывать и дисрегуляцию развития астроцитов и изменять их реактивность. Нарушения нормального развития и созревания мозга должно иметь следствием нарушения межнейрональных контактов и пластичности, связанные с функционированием астроцитов и их отростков, которые могут сказываться на течении болезненного процесса при шизофрении. Все эти данные позволяют предположить важную роль нарушения развития ПФК и дифференцировки астроцитов в патологии этой структуры при шизофрении, возможно приводящие к нарушению взаимоотношений астроцитов с нейронами и к их гипотрофии или атрофии.
Исследование поддержано McDonnell Foundation. Работа была частично выполнена в Университете штата Иллинойс, Урбана-Шампань, США.
Вихрева Ольга Васильевна, кандидат биологических наук, старший научный сотрудник лаборатории клинической нейроморфологии ФГБНУ «Научный центр психического здоровья», Москва, Россия
Olga V. Vikhreva, Cand. of Ski. (Biol.), senior researcher, laboratory of clinical neuromorphology, FSBSI, MHRC, Moscow, Russia
Рахманова Валентина Ивановна, ведущий программист, отдел исследования мозга, лаборатория ультраструктуры и цитохимии мозга, ФГБНУ «Научный центр неврологии», Москва, Россия
Valentina I. Rakhmanova, application programmer, department of brain research, laboratory of the ultrastructure and cytochemistry, FSGSI «Scientific Neurological Center», Moscow, Russia
Клинцова Анна Юрьевна, кандидат биологических наук, профессор факультета психологии и изучения мозга, Университет штата Делавер, США.
Anna Ju. Klintsova, Cand. of Ski. (Biol.), professor, department of psychological and brain sciences, University of Delaware, USA
Гринау Уильям Т., профессор, Университет штата Иллиноис, США
William T. Greenough, PhD, swanland professor, Beckman Institute, University of Illinois at Urbana-Champaign, USA
Уранова Наталия Александровна, доктор медицинских наук, заведующая лабораторией клинической нейроморфологии ФГБНУ «Научный центр психического здоровья», Москва, Россия
Natalya A. Uranova, Dr. of Sci. (Med.), head of the laboratory of clinical neuromorphology, FSBSI, MHRC, Moscow, Russia
Литература References
1. Zhou Y., Fan L., Qiu C., Jiang T. Prefrontal cortex and the dysconnectivity hypothesis of schizophrenia. Neurosci. Bull. 2015;31(2):207-219. doi: 0.1007/s12264-014-1502-8
Zhou Y., Fan L., Qiu C., Jiang T. Prefrontal cortex and the dysconnectivity hypothesis of schizophrenia. Neurosci. Bull. 2015;31(2):207-219. doi: 0.1007/s12264-014-1502-8
2. Sanfilipo M., Lafargue T., Rusinek H., Arena L., Loneragan C., Lautin A., Feiner D., Rotrosen J., Wolkin A. Volumetric measure of the frontal and temporallobe regions in schizophrenia: relationship to negative symptoms. Arch. Gen. Psychiatry. 2000;57:471-480.
Sanfilipo M., Lafargue T., Rusinek H., Arena L., Loneragan C., Lautin A., Feiner D., Rotrosen J., Wolkin A. Volumetric measure of the frontal and temporallobe regions in schizophrenia: relationship to negative symptoms. Arch. Gen. Psychiatry. 2000;57:471-480.
3. Rajkowska G.,Miguel-Hidalgo J.J., Makkos Z., Meltzer H., Overholser J., Stockmeier C. Layer-specific reductions in GFAP-reactive astroglia in the dorsolateral prefrontal cortex in schizophrenia. Schizophr. Res. 2002;57(2-3):127-138.
Rajkowska G.,Miguel-Hidalgo J.J., Makkos Z., Meltzer H., Overholser J., Stockmeier C. Layer-specific reductions in GFAP-reactive astroglia in the dorsolateral prefrontal cortex in schizophrenia. Schizophr. Res. 2002;57(2-3):127-138.
4. Schnieder T.P., Dwork A.J. Searching for neuropathology: gliosis in schizophrenia. Biol. Psychiatry. 2011;69(2):3134-3139. doi: 10.1016/j.biopsych.2010.08.027
Schnieder T.P., Dwork A.J. Searching for neuropathology: gliosis in schizophrenia. Biol. Psychiatry. 2011;69(2):3134-3139. doi: 10.1016/j.biopsych.2010.08.027
5. Востриков В.М., Уранова Н.А., Рахманова В.И., Орловская Д.Д. Сниженная плотность олигодендроглии в префронтальной коре при шизофрении. Журн. неврол. и психиат. им. С.С. Корсакова. 2004;104(1):47-51.
Vostrikov V.M., Uranova N.A., Rakhmanova V.I., Orlovskaia D.D. [Lowered oligodendroglial cell density in the prefrontal cortex in schizophrenia]. Zhurnal nevrologii i psikhiatrii im. S.S. Korsakova. 2004;104(1):47-51. (In Russ.).
6. Vostrikov V., Orlovskaya D., Uranova N. Deficit of pericapillary oligodendrocytes in the prefrontal cortex in schizophrenia. World J. Biol. Psychiatry. 2008;9:34-42.
Vostrikov V., Orlovskaya D., Uranova N. Deficit of pericapillary oligodendrocytes in the prefrontal cortex in schizophrenia. World J. Biol. Psychiatry. 2008;9:34-42.
7. Thune J.J., Uylings H.B., Pakkenberg B. No deficit in total number of neurons in the prefrontal cortex in schizophrenics. J. Psychiatr. Res. 2001;35:15-21.
Thune J.J., Uylings H.B., Pakkenberg B. No deficit in total number of neurons in the prefrontal cortex in schizophrenics. J. Psychiatr. Res. 2001;35:15-21.
8. Black J.E., Kodish I.M., Grossman A.W., Klintsova A.Y., Orlovskaya D., Vostrikov V., Uranova N., Greenough W.T. Pathology oflayer V pyramidal neurons in the prefrontal cortex of patients with schizophrenia. Am. J. Psychiatry. 2004;161:742-744.
Black J.E., Kodish I.M., Grossman A.W., Klintsova A.Y., Orlovskaya D., Vostrikov V., Uranova N., Greenough W.T. Pathology oflayer V pyramidal neurons in the prefrontal cortex of patients with schizophrenia. Am. J. Psychiatry. 2004;161:742-744.
9. Harrison P.J., Weinberger D.R. Schizophrenia genes, gene expression, and neuropathology: on the matter of their convergence. Mol. Psychiatry. 2005;10(1):40-68.
Harrison P.J., Weinberger D.R. Schizophrenia genes, gene expression, and neuropathology: on the matter of their convergence. Mol. Psychiatry. 2005;10(1):40-68.
10. Ellis S.E., Panitch R., West A.B., Arking D.E. Transcriptome analysis of cortical tissue reveals shared sets of downregulated genes in autism and schizophrenia. Transl. Psychiatry. 2016;6:e817. DOI: 10.1038/tp.2016.87
Ellis S.E., Panitch R., West A.B., Arking D.E. Transcriptome analysis of cortical tissue reveals shared sets of downregulated genes in autism and schizophrenia. Transl. Psychiatry. 2016;6:e817. DOI: 10.1038/tp.2016.87
11. Roberts R.C., Conley R., Kung L., Peretti F.J., Chute D.J. Reduced striatal spine size in schizophrenia: a postmortem ultrastructural study. Neuroreport. 1996;7(6):1214-1218.
Roberts R.C., Conley R., Kung L., Peretti F.J., Chute D.J. Reduced striatal spine size in schizophrenia: a postmortem ultrastructural study. Neuroreport. 1996;7(6):1214-1218.
12. Uranova N., Orlovskaya D., Zimina I., Vikhreva O., Rachmanova V., Klintsova A., Black J.E., Grenough W.T. Decreased synaptic size in schizophrenic cortex: a postmortem morphometric electron microscopic study. 30th Annual Meeting of the Society for Neuroscience Abstracts. 2000;26:1558-1558.
Uranova N., Orlovskaya D., Zimina I., Vikhreva O., Rachmanova V., Klintsova A., Black J.E., Grenough W.T. Decreased synaptic size in schizophrenic cortex: a postmortem morphometric electron microscopic study. 30th Annual Meeting of the Society for Neuroscience Abstracts. 2000;26:1558-1558.
13. Uranova N.A., Casanova M.F., DeVaughn N.M., Orlovskaya D.D., Denisov D.V. Ultrastructural alterations of synaptic contacts and astrocytes in postmortem caudate nucleus of schizophrenic patients. Schizophr. Res. 1996;22:81-83.
Uranova N.A., Casanova M.F., DeVaughn N.M., Orlovskaya D. D., Denisov D.V. Ultrastructural alterations of synaptic contacts and astrocytes in postmortem caudate nucleus of schizophrenic patients. Schizophr. Res. 1996;22:81-83.
14. Kung L., Roberts R.C. Mitochondrial pathology in human schizophrenic striatum: a postmortem ultrastructural study. Synapse. 1999;31(1):67-75.
Kung L Roberts R.C. Mitochondrial pathology in human schizophrenic striatum: a postmortem ultrastructural study. Synapse. 1999;31(1):67-75.
15. Somerville S.M., Conley R.R., Roberts R.C. Mitochondria in the striatum of subjects with schizophrenia. World J. Biol. Psychiatry. 2011;12(1):48-56. DOI: 10.3109/15622975.2010.505662
Somerville S.M., Conley R.R., Roberts R.C. Mitochondria in the striatum of subjects with schizophrenia. World J. Biol. Psychiatry. 2011;12(1):48-56. DOI: 10.3109/15622975.2010.505662
16. Roberts R.C., Barksdale K.A., Roche J.K., Lahti A.C. Decreased synaptic and mitochondrial density in the postmortem anterior cingulate cortex in schizophrenia. Schizophr. Res. 2015;168(1-2):543-553. DOI: 10.1016/j. schres.2015.07.01
Roberts R.C., Barksdale K.A., Roche J.K., Lahti A.C. Decreased synaptic and mitochondrial density in the postmortem anterior cingulate cortex in schizophrenia. Schizophr. Res. 2015;168(1-2):543-553. DOI: 10.1016/j.schres.2015.07.01
17. Уранова Н.А., Вихрева О.В., Рахманова В.И., Клинцова А.Ю., Блэк Д., Гринау В.Т., Орловская Д.Д. Нарушения численной плотности синаптических связей в коре больших полушарий при шизофрении. Вестник РАМН. 2007;3:8-14.
Uranova N.A., Vikhreva O.V., Zimina I.S., Rakhmanova V.I., Klintsova A.Iu., Black J., Greenough W.T., Orlovskaia D.D. [Abnormal patterns of cortical synaptic connectivity in schizophrenia]. Vestn. Ross. Akad. Med. Nauk. 2007;3:8-14. (In Russ.).
18. Perez-Alvarez A., Navarrete M., Covelo A., Martin E.D., Araque A. Structural and functional plasticity of astrocyte processes and dendritic spine interactions. J. Neurosci. 2014;34(38):12738-12744. DOI: 10.1523/ JNEUROSCI.2401-14.2014
Perez-Alvarez A., Navarrete M., Covelo A., Martin E.D., Araque A. Structural and functional plasticity of astrocyte processes and dendritic spine interactions. J. Neurosci. 2014;34(38):12738-12744. DOI: 10.1523/JNEUROSCI.2401-14.2014
19. Boksha I.S., Tereshkina E.B., Burbaeva G.Sh. Glutamine synthetase and glutamine synthetase-like protein from human brain, purification and comparative characterization. J. Neurochem. 2000;75:2574-2582.
Boksha I.S., Tereshkina E.B., Burbaeva G.Sh. Glutamine synthetase and glutamine synthetase-like protein from human brain, purification and comparative characterization. J. Neurochem. 2000;75:2574-2582.
20. Burbaeva G.S, Boksha I.S., Tereshkina E.B., Savushkina O.K., Starodubtseva L.I., Turishcheva M.S., Mukaetova-Ladinska E. Systemic neurochemical alterations in schizophrenic brain, glutamate metabolism in focus. Neurochem. Res. 2007;32(9):1434-1444. doi.org/10.1007/ s11064-007-9328-7
Burbaeva G.S, Boksha I.S., Tereshkina E.B., Savushkina O.K., Starodubtseva L.I., Turishcheva M.S., Mukaetova-Ladinska E. Systemic neurochemical alterations in schizophrenic brain, glutamate metabolism in focus. Neurochem. Res. 2007;32(9):1434-1444. doi.org/10.1007/s11064-007-9328-7
21. Xia M., Abazyan A., Yan Jouroukhin Y., Pletnikov M. Behavioral sequelae of astrocyte dysfunction: focus on animal models of schizophrenia. Schizophr Res. 2016;176(1):72-82. DOI: 10.1016/j.schres.2014.10.044
Xia M., Abazyan A., Yan Jouroukhin Y., Pletnikov M. Behavioral sequelae of astrocyte dysfunction: focus on animal models of schizophrenia. Schizophr Res. 2016;176(1):72-82. DOI: 10.1016/j.schres.2014.10.044
22. Webster M., Knable M., Johnston-Wilson N., Nagata K., Inagaki M., Yolken R. Immunohistochemicallocalization of phosphorylated glial fibrillary acidic protein in the prefrontal cortex and hippocampus from patients with schizophrenia, bipolar disorder, and depression. Brain Behav. Immunol. 2001;15:388-400.
Webster M., Knable M., Johnston-Wilson N., Nagata K., Inagaki M., Yolken R. Immunohistochemicallocalization of phosphorylated glial fibrillary acidic protein in the prefrontal cortex and hippocampus from patients with schizophrenia, bipolar disorder, and depression. Brain Behav. Immunol. 2001;15:388-400.
23. Steffek A.E., McCullumsmith R.E., Haroutunian V., Meador-Woodruff J.H. Cortical expression of glial fibrillary acidic protein and glutamine synthetase is decreased in schizophrenia. Schizophr. Res. 2008;103:71-82.
Steffek A.E., McCullumsmith R.E., Haroutunian V., Meador-Woodruff J.H. Cortical expression of glial fibrillary acidic protein and glutamine synthetase is decreased in schizophrenia. Schizophr. Res. 2008;103:71-82.
24. Feresten A.H., Barakauskas V., Ypsilanti A., Barr A.M., Beasley C.L Increased expression of glial fibrillary acidic protein in prefrontal cortex in psychotic illness. Schizophr. Res. 2013;150:252-257.
Feresten A.H., Barakauskas V., Ypsilanti A., Barr A.M., Beasley C.L. Increased expression of glial fibrillary acidic protein in prefrontal cortex in psychotic illness. Schizophr. Res. 2013;150:252-257.
25. Catts V.S., Wong J., Fillman S.G., Fung S.J., Weickert S.C. Increased expression of astrocyte markers in schizophrenia: Association with neuroinflammation. Aust. NZ. J. Psychiatry. 2014;48(8):722 - 734. DOI: 10.1177/0004 8 67414531078
Catts V.S., Wong J., Fillman S.G., Fung S.J., Weickert S.C. Increased expression of astrocyte markers in schizophrenia: Association with neuroinflammation. Aust. NZ. J. Psychiatry. 2014;48(8):722 - 734. DOI: 10.1177/0004867414531078
26. Kolomeets N.S., Uranova N. Ultrastructural abnormalities of astrocytes in the hippocampus in schizophrenia and duration of illness: a postmortem morphometric study. World J. Biol. Psychiatry. 2010;11(2):282-292.
Kolomeets N.S., Uranova N. Ultrastructural abnormalities of astrocytes in the hippocampus in schizophrenia and duration of illness: a postmortem morphometric study. World J. Biol. Psychiatry. 2010;11(2):282-292.
27. Blanco-Su rez E., Caldwell A.L., Allen N.J. Role of astrocyte-synapse interactions in CNS disorders. J. Physiol. 2017;595(6):1903-1916. DOI: 10.1113/JP270988
Blanco-Su rez E., Caldwell A.L., Allen N.J. Role of astrocyte-synapse interactions in CNS disorders. J. Physiol. 2017;595(6):1903-1916. DOI: 10.1113/JP270988
28. Sirevaag A.M., Greenough WT. Plasticity of GFAP-immunoreactive astrocyte size and number in visual cortex of rats reared in complex environments. Brain Res. 1991;540(1-2):273-278.
Sirevaag A.M., Greenough WT. Plasticity of GFAP-immunoreactive astrocyte size and number in visual cortex of rats reared in complex environments. Brain Res. 1991;540(1-2):273-278.
29. Greenough W.T., Chang F.L. Dendritic pattern formation involves both oriented regression and oriented growth in the barrels of mouse somatosensory cortex. Brain Res. 1988;471(1):148-152.
Greenough W.T., Chang F.L. Dendritic pattern formation involves both oriented regression and oriented growth in the barrels of mouse somatosensory cortex. Brain Res. 1988;471(1):148-152.
30. Ойфа А.И., Уранова Н.А. Электронномикроскопический анализ цитоархитектонических нарушений в коре мозга при шизофрении. Журнал неврологии и психиатрии им. С.С. Корсакова. 1991;91(10):48-52.
O fa A.I., Uranova N.A. Electron-microscopic analysis of cytoarchitectonic disorders in the cerebral cortex in schizophrenia. Zhurnal neurologii i psikhiatrii im. S.S. Korsakova. 1991;91(10):48-52. (In Russ.).
31. Davis J.M. Dose equivalence of the antipsychotic drugs. J. Psychiat. Res. 1974;11:65-69.
Davis J.M. Dose equivalence of the antipsychotic drugs. J. Psychiat. Res. 1974;11:65-69.
32. Woods S.W. Chlorpromazine equivalent doses for the new atypical antipsychotics. J. Clin. Psychiatry. 2003;64(6):663-667.
Woods S.W. Chlorpromazine equivalent doses for the new atypical antipsychotics. J. Clin. Psychiatry. 2003;64(6):663-667.
33. Питерс А., Палей С., Уэбстер Г. Ультраструктура нервной системы. Пер. с англ. М.: Мир, 1972.
Peters A, Paley C, Webster G. The fine structure of the nervous system. 1976. W.B. Saunders Company
34. Gundersen H.J., Bagger P., Bendtsen T.F., Evans S.M., Korbo L., Marcussen N., M ller A., Nielsen K., Nyengaard J.R., Pakkenberg B. The new stereological tools: disector, fractionator, nucleator and point sampled intercepts and their use in pathological research and diagnosis. APMIS. 1988;96:857-881.
Gundersen H.J., Bagger P., Bendtsen T.F., Evans S.M., Korbo L., Marcussen N., M ller A., Nielsen K., Nyengaard J.R., Pakkenberg B. The new stereological tools: disector, fractionator, nucleator and point sampled intercepts and their use in pathological research and diagnosis. APMIS. 1988;96:857-881.
35-.Konopaske G.T., Dorph-Petersen K.A., Pierri .J.N., Wu Q., Sampson A.R., Lewis D.A. Effect of chronic exposure to antipsychotic medication on cell numbers in the parietal cortex of macaque monkeys. Neuropsychopharmacol. 2007;32:1216-1223.
Konopaske G.T., Dorph-Petersen K.A., Pierri .J.N., Wu Q., Sampson A.R., Lewis D.A. Effect of chronic exposure to antipsychotic medication on cell numbers in the parietal cortex of macaque monkeys. Neuropsychopharmacol. 2007;32:1216-1223.
36. Коломеец Н.С., Востриков В.М., Уранова Н.А. Влияние сыворотки крови больных шизофренией на монотерапии оланзапином на ультраструктуру астроцитов в органотипической культуре ткани эмбрионального мозга человека. Журнал неврологии и психиатрии им. С.С. Корсакова. 2016;116(5):65-70. DOI: 10.17116/ jnevro20161165165-70
Kolomeets N.S., Vostrikov V.M., Uranova N.A. The effects of blood serum from schizophrenia patients under olanzapine monotherapy on the ultrastructure of astrocytes in human fetal brain organotypic culture. Zhurnal neurologii i psikhiatrii im. S.S. Korsakova. 2016;116(5):65-70. DOI: 10.17116/ jnevro20161165165-70
37. Sullivan C.R., O'Donovan S.M., McCullumsmith R.E., Ramsey A. Defects in Bioenergetic Coupling in Schizophrenia. Biol. Psychiatry. 2017 pii: S0006-3223(17)32099-1. DOI: 10.1016/j.biopsych.2017.10.014
Sullivan C.R., O'Donovan S.M., McCullumsmith R.E., Ramsey A. Defects in Bioenergetic Coupling in Schizophrenia. Biol. Psychiatry. 2017 pii: S0006-3223(17)32099-1. DOI: 10.1016/j. biopsych.2017.10.014
38. Rezin G.T., Amboni G., Zugno A.I., Quevedo J., Streck E.L. Mitochondrial dysfunction and psychiatric disorders. Neurochem. Res. 2009;34(6):1021-1029. DOI: 10.1007/ s11064-008-9865-8
Rezin G.T., Amboni G., Zugno A.I., Quevedo J., Streck E.L. Mitochondrial dysfunction and psychiatric disorders. Neurochem. Res. 2009;34(6):1021-1029. DOI: 10.1007/s11064-008-9865-8
39. Ben-Shachar D., Karry R. Neuroanatomical pattern of mitochondrial complex I pathology varies between schizophrenia, bipolar disorder and major depression. PLoS One. 2008;3(11):e3676. DOI: 10.1371/journal.pone.0003676
Ben-Shachar D., Karry R. Neuroanatomical pattern of mitochondrial complex I pathology varies between schizophrenia, bipolar disorder and major depression. PLoS One. 2008;3(11):e3676. DOI: 10.1371/journal.pone.0003676
40. Martins-de-Souza D., Harris L.W., Guest P.C., Bahn S. The role of energy metabolism dysfunction and oxidative stress in schizophrenia revealed by proteomics. Antioxid Redox Signal. 2011;15(7):2067-2079. DOI: 10.1089/ ars.2010.3459
Martins-de-Souza D., Harris L.W., Guest P.C., Bahn S. The role of energy metabolism dysfunction and oxidative stress in schizophrenia revealed by proteomics. Antioxid Redox Signal. 2011;15(7):2067-2079. DOI: 10.1089/ars.2010.3459
41. van de Ven V., Rotarska Jagiela A., Oertel-Kn chel V., Linden D.E.J. Reduced intrinsic visual cortical connectivity is associated with impaired perceptual closure in schizophrenia. Neuroimage Clin. 2017;15:45-52. DOI: 10.1016/j.nicl.2017.04.012
van de Ven V., Rotarska Jagiela A., Oertel-Kn chel V., Linden D. E.J. Reduced intrinsic visual cortical connectivity is associated with impaired perceptual closure in schizophrenia. Neuroimage Clin. 2017;15:45-52. DOI: 10.1016/j. nicl.2017.04.012
42. Lang X., Wang L., Zhuo C.J., Jia F., Wang L.N., Wang C.L. Reduction of Interhemispheric Functional Connectivity in Sensorimotor and Visual Information Processing Pathways in Schizophrenia. Chin. Med. J. (Engl) 2016;129(20):2422-2426. DOI: 10.4103/0366-6999.191758
Lang X., Wang L., Zhuo C.J., Jia F., Wang L.N., Wang C.L. Reduction of Interhemispheric Functional Connectivity in Sensorimotor and Visual Information Processing Pathways in Schizophrenia. Chin. Med. J. (Engl) 2016;129(20):2422-2426. DOI: 10.4103/0366-6999.191758
43. Muly E.C., Maddox M., Smith Y. Distribution of mGluR1alpha and mGluR5 immunolabeling in primate prefrontal cortex. J. Comp. Neurol. 2003;467(4):521-535.
Muly E.C., Maddox M., Smith Y. Distribution of mGluR1alpha and mGluR5 immunolabeling in primate prefrontal cortex. J. Comp. Neurol. 2003;467(4):521-535.
44. Roberts R.C., Roche J.K., McCullumsmith R.E. Localization of excitatory amino acid transporters EAAT1 and EAAT2 in human postmortem cortex: alight and electron microscopic study. Neuroscience. 2014;277:522-540.
Roberts R.C., Roche J.K., McCullumsmith R.E. Localization of excitatory amino acid transporters EAAT1 and EAAT2 in human postmortem cortex: alight and electron microscopic study. Neuroscience. 2014;277:522-540.
45. Dracheva S., McGurc S.R., Haroutunian V. mRNA expression of AMPA receptors and AMPA receptor binding proteins in the cerebral cortex of elderly schizophrenics. J. Neurosci. Res. 2005;79(6):868-878.
Dracheva S., McGurc S.R., Haroutunian V. mRNA expression of AMPA-receptors and AMPA-receptor binding proteins in the cerebral cortex of elderly schizophrenics. J. Neurosci. Res. 2005;79(6):868-878.
46. van Elst L.T., Valerius G., Buchert M., Thiel T., R sch N., Bubl E., Hennig J., Ebert D., Olbrich H.M. Increased prefrontal and hippocampal glutamate concentration in schizophrenia: evidence from a magnetic resonance spectroscopy study. Biol. Psychiatry. 2005;58(9):724- 730.
van Elst L.T., Valerius G., Buchert M., Thiel T., R sch N., Bubl E. , Hennig J., Ebert D., Olbrich H.M. Increased prefrontal and hippocampal glutamate concentration in schizophrenia: evidence from a magnetic resonance spectroscopy study. Biol. Psychiatry. 2005;58(9):724-730.
47. Scarr E., Beneyto M., Meador-Woodruff J.H., Deans B. Cortical glutamatergic markers in schizophrenia. Neuropsychopharmacology. 2005;30(8):1521-1531.
Scarr E., Beneyto M., Meador-Woodruff J.H., Deans B. Cortical glutamatergic markers in schizophrenia. Neuropsychopharmacology. 2005;30(8):1521-1531.
48. Goudriaan A., de Leeuw C., Ripke S., Hultman C.M., Sklar P., Sullivan P.F., Smit A.B., Posthuma D., Verheijen M.H. Specific Glial Functions Contribute to Schizophrenia Susceptibility. Schizophr. Bull. 2014;40:925-935.
Goudriaan A., de Leeuw C., Ripke S., Hultman C.M., Sklar P., Sullivan P.F., Smit A.B., Posthuma D., Verheijen M.H. Specific Glial Functions Contribute to Schizophrenia Susceptibility. Schizophr. Bull. 2014;40:925-935.
49. Hagihara H., Ohira K., Takao K., Miyakawa T. Transcriptomic evidence for immaturity of the prefrontal cortex in patients with schizophrenia. Mol. Brain. 2014;7:41-41.
Hagihara H., Ohira K., Takao K., Miyakawa T. Transcriptomic evidence for immaturity of the prefrontal cortex in patients with schizophrenia. Mol. Brain. 2014;7:41-41.
50. Markham J.A., Mullins S.E., Koenig J.I. Periadolescent maturation of the prefrontal cortex is sex-specific and is disrupted by prenatal stress. J. Comp. Neurol. 2013;521:1828-1843.
Markham J.A., Mullins S.E., Koenig J.I. Periadolescent maturation of the prefrontal cortex is sex-specific and is disrupted by prenatal stress. J. Comp. Neurol. 2013;521:1828-1843.



