Механизмы воздействия магния и пиридоксина на структуру и свойства соединительной ткани как основание для магнезиальной терапии дисплазий соединительной ткани
СтатьиО.А. Громова1, И.Ю. Торшин1, Е.Ю. Егорова1,2
2- ГОУ ВПО Ивановский государственный университет
Соединительная ткань - структурная поддержка, своего рода «экзоскелет» любой ткани организма, в любом возрасте. Соединительная ткань начинает развиваться в эмбриональном периоде из мезинхимы и в, последующем, формирует опорный каркас (фасции, связки и костную ткань), кожные покровы ребенка. Поэтому, нарушения структуры соединительной ткани, обусловленные нутрициальными и генетическими факторами, будут способствовать развитию многочисленных заболеваний у детей. Эти нарушения, известные также как недифференцированная дисплазия соединительной ткани (нДСТ), особенно опасны в детском возрасте, когда происходит интенсивный рост всех тканей организма. Как показывают результаты фундаментальных и клинических исследований, обеспеченность и магнием, и пиридоксином важна для нормального роста и поддержания физиологического состояния тканей в процессе роста и развития детского организма. В настоящей статье результаты проведенного нами анализа молекулярных механизмов воздействия магния на структуру соединительной ткани. Использование фенотипических маркеров нДСТ с учетом разбираемых механизмов позволяет осуществлять рациональную терапию нДСТ посредством современных препаратов магния.
Molecular mechanisms of magnesium and pyridoxine in the structure of connective tissue: the basis for magnesium therapy of connective tissue dysplasias
OA Gromova, IY Torshin, EY Egorova
Russian Satellite Centre of the Trace Element Institute for UNESCO, Moscow,
Laboratory for Computational and Systems Biology, Computing Center RAS, Moscow
Connective tissue is a structural support, a sort of "exoskeleton" for any tissue of the body. In embryonic period, the connective tissue starts to develop from mesenchyme and, subsequently, differentiates into fascia, ligaments, bone and the skin. Apparentlly, any disruption of the structure of connective tissue, caused either by nutritional or genetic factors, will contribute to the development of numerous diseases in children. These disruptions, collectively called as “undifferentiated connective tissue dysplasia” (uCTD), are especially dangerous in childhood, the period of the rapid growth of all body tissues. As indicate the results of basic and clinical research, adequate nutritional support with magnesium and pyridoxine is important for normal growth and maintenance of the physiological state of the connective tissue. In this paper we present the results of analysis of the molecular mechanisms of magnesium and pyridoxine influence on the structure of connective tissue. Using phenotypic markers of uCTD along with the information on these molecular mechanisms allows to conduct rational therapy of uCTD by modern pharmaceutical forms of magnesium.
Keywords: magnesium deficit, connecting tissue dysplasia, extracellular matrix, bioinformatics.
Введение
В аналитическом отчете академика РАМН Баранова А.А. (2010) указано, что в последние годы (2000-2010) педиатры отмечают уменьшение детей относящихся к 1 группе здоровья и увеличивающуюся инвалидизацию детского населения России [1]. Эти отклонения прямо указывают на несовершенство существующих профилактических стратегий и, во многом, обусловлены неполноценным развитием соединительной ткани, называемым «дефицитом соединительной ткани» или «недифференцированной дисплазией соединительной ткани» (нДСТ).
Дисплазия соединительной ткани – нутрициально и генетически детерминированное состояние, обусловленное нарушениями метаболизма соединительной ткани в эмбриональном и постнатальном периодах и характеризующееся аномалиями структуры компонентов внеклеточного матрикса (волокон и основного вещества гелеобразной среды) с прогредиентными морфо-функциональными изменениями различных систем и органов [2].
Следует отметить очевидность широкого популяционного распространения ДСТ у детей и подростков. Недифференцированная ДСТ встречается в России гораздо чаще: 20% в когорте около 3000 подростков и лиц молодого возраста (по данным проф. Нечаевой Г.И., 1980-2010) [2]; 8,5% в выборке из 400 человек [3]. Данных распространенности нДСТ по другим странам не имеется и российские исследователи являются лидерами в области диагностики, фундаментальных молекулярных исследований и терапии нДСТ.
На сегодняшний день установлено, что диспластико-зависимые морфо-функциональные изменения систем органов существенным образом сказываются на течение сопутствующих заболеваний, определяя затяжное течение и хронизацию острых процессов, меньшую эффективность традиционных схем лечения, более длительный период реконвалисценции и т.д. [2,4].
Значение магния в педиатрической практике трудно переоценить. В настоящее время в педиатрии наблюдается подъем исследовательского интереса к клиническим приложениям магния. Терапевтические роли магния (Mg) в лечении сердечно-сосудистых, обменных, мышечно-тетанических и неврологических нарушений у детей основаны на фундаментальной роли, которую играет гомеостаз магния в организме человека [5,6]. Как показали результаты проведенного нами систематического анализа взаимодействий между магнием и метаболизмом соединительной ткани, на фоне дефицита магния диспластические процессы в соединительной ткани будут неизбежно обострятся [7]. В настоящей работе подробно рассматриваются основы диагностики нДСТ, молекулярно-биологические механизмы обеспечивающие функционирование соединительной ткани и роли магния с пиридоксином в поддержке этих молекулярных механизмов.
Клинико-морфологические особенности ДСТ
В зависимости от особенностей этиологического фактора выделяют дифференцированные и недифференцированные формы ДСТ. К дифференцированным (синдромным) ДСТ относят болезни монофакторного характера с установленным генным дефектом, известным типом наследования и, как правило, с выраженной и четко очерченной клинической симптоматикой. Классическим примером синдромных ДСТ являются синдромы Марфана и Элерса-Данло, несовершенный остеогенез и некоторые другие редкие генетические синдромы.
Недифференцированные формы ДСТ имеют полигенно-мультифакториальную природу, т.е. в их возникновении играют роль как полиморфизмы или мутации большого числа генов в различных сочетаниях, так и воздействие разнообразных факторов внешней среды. Клинические проявления недифференцированной ДСТ не укладываются ни в одну из известных дифференцированных наследственных болезней, хотя иногда могут их напоминать. Принято выделять марфаноидный, элерсоподобный и MASS-подобный фенотип (по первым буквам наиболее частых фенотипических признаков ДСТ – англ. «Mitral valve», «Aorta», «Skeleton», «Skin»), предполагая единую генетическую сущность данных состояний.
Имеющиеся на настоящий момент данные свидетельствуют, что частота встречаемости ДСТ зависит от возраста обследованных лиц. Признаки нДСТ проявляются в течение жизни: в период новорожденности выявление признаков нДСТ минимально; в возрасте 4-5 лет начинают формироваться пролапсы клапанов сердца, в 10-12 лет – торако-диафрагмальный синдром (деформации грудной клетки и позвоночника), плоскостопие, миопия, в подростковом и молодом возрасте - сосудистый синдром [8].
Критическим периодом является подростковый возраст, когда прирост количества разбираемых ниже признаков дисморфогенеза соединительной ткани максимально [9].
Фенотипические проявления ДСТ условно можно разделить на группы в зависимости от органов и систем, вовлеченных в диспластический процесс. Ниже, приводятся основные морфологические признаки нДСТ [2-8]. Хотя перечисленные ниже признаки, будучи отдельно взятыми, не являются строго специфичными для ДСТ, анализ совокупности этих признаков является основой диагностики ДСТ у ребенка.
Костно-суставные изменения
Астенический тип конституции
Долихостеномелия
Арахнодактилия
Деформации грудной клетки (воронкообразные и килевидные)
Деформации позвоночника (сколиоз, синдром прямой спины, гиперкифоз, гиперлордоз, спондилолистез)
Деформации черепа (акроцефалия, арковидное небо, микрогнатия, скученность зубов)
Деформации конечностей (вальгусная, варусная)
Деформации стопы (плоскостопие, полая стопа и др.)
Гипермобильность суставов
Изменения кожи и мышц
Растяжимая кожа
Тонкая кожа
Вялая кожа
Заживление в виде «папиросной бумаги»
Келлоидные рубцы
Геморрагические проявления (экхимозы, петехии)
Мышечная гипотония и/или гипотрофия
Грыжи
Признаки ДСТ органа зрения
Миопия
Плоская роговица
Подвывих (вывих) хрусталика
Признаки ДСТ сердечно-сосудистой системы
Пролапсы клапанов
Миксоматозная дегенерация клапанных структур
Дилятация фиброзных колец
Расширение корня аорты
Аневризмы межпредсердной, межжелудочковой перегородки
Расширение и аневризмы сосудов (аорта, легочная артерия, церебральные артерии)
Варикозное расширение вен, флебопатии
Нарушения сосудистого гемостаза, тромбоцитопатии,
Признаки ДСТ бронхолегочной системы
Трахеобронхомаляция, трахеобронхомегалия
Трахеобронхиальная дискинезия
Бронхоэктазы
Апикальные буллы и первичный спонтанный пневмоторакс
Признаки ДСТ пищеварительной системы
Моторно-тонические нарушения (рефлюксы)
Нарушения фиксации органов (гастроптоз, колоноптоз)
Изменения размеров и длины полых органов (мегаколон, долихосигма и др.)
Признаки ДСТ мочевыделительной системы
Нефроптоз, рефлюксы
Особенностью диспластического фенотипа является частое сочетание ДСТ с пороками развития и/или малыми аномалиями развития (МАР) со стороны различных органов и систем. Попытки расценить порочное развитие органов или малые аномалии развития как признаки ДСТ приводят к гипердиагностике ДСТ и дезориентируют исследователей и практических врачей (табл. 1).
Таблица 1.
Малые аномалии развитияу детей [10].
| Голова: | Область рта и ротовой полости: |
| Аномальный рисунок роста волос | Расщепление язычка |
| Орбительная область: | Аберрантные уздечки преддверия рта |
| Эпикантные складки | Микродентия |
| Монголоидный разрез глаз | Аномально растущие зубы |
| Антимонголоидный разрез глаз | Шея: |
| Короткие глазные щели | Крыловидная шея |
| Дистопия наружных углов глаза | Кисти: |
| Гипотелоризм умеренный | Рудиментарная полидактилия |
| Гипертелоризм умеренный | Аномальная дерматоглифика |
| Птоз легкий | Клинодактилия мизинцев |
| Ушные раковины: | Укорочение 4—5 пальцев |
| Примитивная форма | Гипоплазия терминальных фаланг |
| Дарвинов бугорок | Стопы: |
| Аномальная форма завитка | Синдактилия 2—3 пальцев |
| Оттопыренные | Сандалевидные щели |
| Отсутствие козелка | Короткий 1 палец |
| Расщепление мочки | Наложение пальцев (4-5) |
| Отсутствие мочки | Утолщенные ногти |
| Аурикулярные ямки | Кожные покровы: |
| Нос и фильтр: | Гиперпигментации кожи и невусы |
| Гипоплазия крыльев носа | Монголоидные пятна (у белой расы) |
| Уплощенный фильтр | Депигментации кожи |
| Выступающий фильтр | Добавочные соски или ареолы |
В настоящее, в номенклатуре болезней ВОЗ недифференцированные варианты ДСТ еще не выделены в отдельную рубрику, что несомненно затрудняет работу практического врача. Однако, при внимательной работе с классификацией МКБ можно найти соответствующий код для любого проявления ДСТ (I 34.1 Пролапс митрального клапана, I 71.2 Аневризма и расслоение аорты, I 83 Варикозное расширение вен нижних конечностей, М35.7 Гипермобильный синдром и многие другие).
Диагностика ДСТ и последующее обследование ребенка включает следующие этапы: сбор анамнеза; физикальное и лабораторно-инструментальные обследование; генеалогический анамнез. Тщательно собранный анамнез обеспечивает получение важной информации о манифестации, темпах прогрессирования ДСТ, наличии факторов риска, признаках поражения того или иного органа или системы.
Обследование детей и подростков с ДСТ проводятся в строгой последовательности, в соответствии со следующими задачами:
Поиск фенотипических признаков ДСТ должен проводиться при физикальном обследовании целенаправленно и последовательно. Наиболее подробно методы диагностики нДСТ описаны в монографии Нечаевой Г.И. с соавт (2009) [2].
Молекулярная биология соединительной ткани
Диагностика нДСТ на основе разобранных выше клинических показателей является показанием для терапии. В настоящем разделе представлен краткий обзор данного вопроса, более подробная информация приводится в работе [11].
Составляя около 50% массы тела, соединительная ткань (СТ) является одним из четырех основных типов ткани в традиционных классификациях (в дополнение к эпителиальной, мышечной и нервной ткани). Основная функция СТ - это структурная поддержка, своего рода «экзоскелет» для всех других тканей организма. Хрящ и кость являются основными разновидностями соединительной ткани, другие типы включают ареолярную соединительную ткань, скрепляющую органы и плотную соединительную ткани, формирующую связки и сухожилия.
В отличие от эпителия, в котором клетки плотно связаны вместе посредством механизмов межклеточной адгезии посредством интегринов и кадхеринов, соединительная ткань демонстрирует избыток внеклеточного матрикса (ВКМ) при достаточно небольшом числе клеток. Именно ВКМ (протеогликаны, коллагены, и эластин) помогает держать клетки и ткани вместе [12]. Только матрикс обеспечивает организованную среду, в пределах которой мигрирующие клетки могут перемещаться и взаимодействовать друг с другом. ВКМ состоит из принципиально необходимых компонентов: гелеобразной среды и волокон клеток (фибробластов, остеобластов или хондробластов, в зависимости от конкретного типа соединительной ткани).
В детском возрасте в соединительной ткани отмечается повышенное содержание гелеобразной среды, в то время как коллагеновых волокон несколько меньше. Характерным для соединительной ткани у детей также является более высокое насыщение водной фракцией, что как раз и обусловлено смещением пропорции «гелеобразная среда-волокна» в сторону гелеобразной среды. Оба эти фактора обуславливают повышенную пластичность всех типов соединительной ткани в детском и подростковом возрасте.
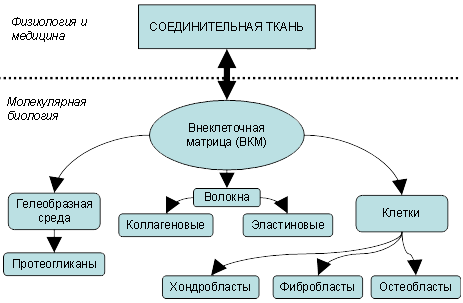
Рис. 1. Структурные компоненты соединительной ткани.
Гелеобразная среда. Наиважнейший компонент внеклеточного матрикса - гелеобразная среда, формируемая протеогликанами, чрезвычайно растянутыми полипептидными цепями с многочисленными полисахаридными цепями глюкозаминогликанов, присоединенных посредством ковалентных связей (Рис. 2.).
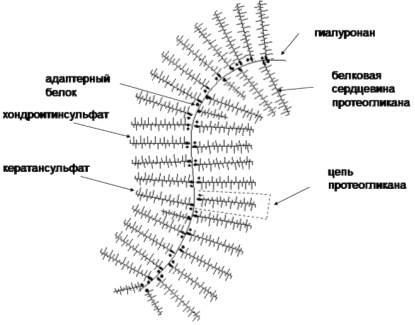
Рис. 2. Основные компоненты протеогликанов - основы гелеобразной среды соединительной ткани.
Многочисленные цепи протеогликанов прикрепляются к особому виду глюкозаминогликана - полимеру гиалуроновой кислоты называемому гиалуронаном. Нити гиалуронана скрепляют структуру геля в единое целое и этот полисахаридный "гель" может противостоять сжатию и растяжению ВКМ и, в то же время, обеспечивает быструю диффузию питательных веществ, строительных материалов и гормонов между кровью и клетками соединительной ткани. Гиалоуронан синтезируется посредством гиалоуронансинтетаз (гены HAS1, HAS2 и HAS3) и деградируется посредством гиалуронидаз (гены HYAL2, HYAL3, HYAL4 и HYALP).
Следует отметить, что ферменты, участвующие в биохимических модификациях и присоединении глюкозаминогликанов, могут значительно влиять на структуру ВКМ. Например, редкие генетические дефекты ксилозил бета-1,4-галактозилтрансферазы 7 (ген B4GALT7) связаны с одной из форм дифференцированной ДСТ - синдрома Элерса-Данло [13], который проявляется как гипермобильность суставов, хрупкая или гиперэластичная кожа, хрупкие кровяные сосуды и т.д. В то же время, пониженная активность этих ферментов вследствие дефицитов необходимых кофакторов таких как магний, медь, цинк, пиридоксин и др. будет способствовать развитию недифференцированной ДСТ. Поэтому, обеспеченность этими микронутриентами особенно важна в детском возрасте, когда темпы формирования соединительной ткани максимальны.
Коллагеновые волокна придают соединительной ткани уже с детства прочность и долговечность. Коллагены - одни из наиболее обильных белков во внеклеточном матриксе и в соединительной ткани. Плотность коллагеновых волокон растет с возрастом и выходит на пик в подростковом и молодом возрасте и жесткость соединительной ткани увеличивается. Именно поэтому для занятий художественной и спортивной гимнастикой, в акробатикой и другими видами спорта требующими высокой гибкости отбираются дети 3 – 8 лет.
Унаследованные мутации в генах коллагенов приводят, в большинстве случаев, к дисплазиям типа Элерса-Данло. Однако,коллагеные болезни чаще всего возникают не столько из-за генетических дефектов в коллагенах, сколько вследствие дефектов в десятках генов, которые влияют на биосинтез, посттрансляционные модификации, секрецию, самосборку и ремоделирование коллагенов. Дефицит активности гена LOX обнаруживался у пациентов с синдромом Элерса-Данло [14].
Деградация (ремоделирование) коллагеновых волокон производится посредством матриксных металлопротеиназ (ММП). Специфические ММП, которые деградируют коллагеновые волокна, таким образом, удаляя основные структурные опоры соединительней ткани известны под названием коллагенaз. Сверхизбыточная активность матриксных металлопротеиназ сопутствует сверхраннему старению в детском возрасте (прогерии, средняя длительность жизни – 13 лет). Прогерия является следствием генетических дефектов в ламине А (ген LMNA), который формирует ламины оболочки клеточного ядра.
Клетки (фибробласты, хондробласты, остеобласты) – активный компонент внеклеточного матрикса и соединительной ткани. Именно клетки ВКМ синтезируют и поддерживают структурную целостность всех вышерасмотренных компонентов соединительной ткани – протеогликанов гелеобразной среды, коллагеновых и эластиновых волокон. С возрастом, профиль активности фибробластов изменяется в сторону поддержки большей плотности коллагеновых волокон.
Магний и структура соединительной ткани
Наиболее общий эффект воздействия Mg2+ на любую ткань заключается в том, что ионы Mg2+ необходимы для стабилизации некодирующих РНК. В частности, ион Mg2+ стабилизируют структуру транспортной РНК и дефицит магния приведет к увеличению числа дисфункциональных молекул тРНК, таким образом, снижая и замедляя общую скорость белкового синтеза.
Совместное рассмотрение молекулярной биологии внеклеточного матрикса и физиологических механизмов гомеостаза магния может позволить нам сформулировать более специфические молекулярные механизмы через которые может осуществляться взаимосвязь ДСТ и дефицита магния. Для того, чтобы найти подобного рода взаимосвязи на уровне индивидуальных молекулярных генов, необходимо рассмотреть каждый из основных элементов ВКМ в отдельности.
Например, гиалоуронансинтетазы HAS1, HAS2 и HAS3 (которые, как мы упомянули ранее, синтезируют основу гелеобразной среды – гиалоуронан) содержат ион магния в активном центре (Рис. 3). С другой стороны известно, что действие ингибиторов гиалуронидаз (ферментов, деградирующих гиалоуронан) также зависит от концентрации ионов магния [16]. Дефицит магния будет приводить к понижению активности гиалоуронансинтетаз и, в то же время, к повышению активности гиалуронидаз (так как ингибиторы перестают действовать при недостатке магния). Оба этих процесса приведут к ухудшению механических свойств нитей гиалоуронана и частичной деградации гелеобразной среды образующей ВКМ. Поэтому, дефицит магния в детском возрасте будет приводить к недостатку гелеобразной среды и, как следствие, ухудшится качество всех соединительно-тканных прослоек организма.
Непосредственно, ионы Mg2+ не взаимодействуют ни с молекулами коллагена, ни с TIMP белками, так что эффект Mg2+ на активность коллагеназ особенно интересен. Известно, что лечение пациентов с острым инфарктом миокарда посредством сульфата магния и оротата магния тормозит повреждение миокарда. Обычно, максимальные уровни IL6 и MMP1 в крови значительно возрастают при остром инфаркте миокарда, но эти уровни остаются на сравнительно низком уровне у пациентов после магний-терапии. Возрастающая концентрация IL6 может приводить к повышению общей активности MMP1, ведя к повреждению тканей, в то время как увеличение концентрации Mg2+ в сыворотке крови уменьшает уровни IL6 и MMP1 [17].
Трансглутаминаза, фермент формирующий поперечные глутамин-лизиновые сшивки соединяющие вместе цепи эластина, активизируется Ca2+ и ингибируется Mg2+ [18]. Mg2+ также может ингибировать медь-зависимую лизилоксидазу (LOX, Рис. 4) [19], также вовлеченную в поперечную сшивку цепей эластинов и/или коллагенов. Соответственно, дефицит Mg2+ может приводить к активизации поперечной сшивки коллагенов и эластинов и этот процесс, наряду с увеличением активности ММП, приведет к своего рода грануляризации соединительной ткани.
Таким образом, имеющиеся данные позволяют сделать вывод, что наиболее вероятные механизмы воздействия дефицита Mg2+ на соединительную ткань - это усиление деградации коллагеновых и, возможно, эластиновых волокон, а также полисахаридных нитей гиалоуронана(Рис. 5). Нельзя исключить, что усиление поперечных сшивок приведет к грануляризации соединительной ткани, расслоению на этакие «пластинки» состоящие из наполовину деградированных молекул коллагенов и, в результате, приводит к уменьшению механической прочности. При достаточной концентрации Mg2+ секреция/активность ММП снижаются, что приводит к уменьшению деградации и ускорению белкового синтеза новых молекул коллагенов [11-19].
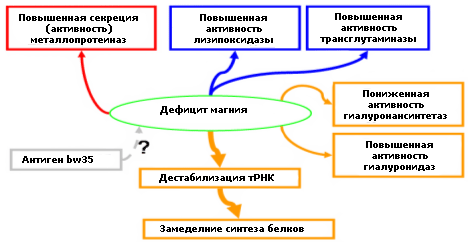
Рис 5. Предлагаемые механизмы, связывающие дефицит магния со структурой соединительной ткани
Пиридоксин и соединительная ткань
Пиридоксин – синергист магния, способствующий более эффективному всасыванию минерала. Помимо этого, пиридоксин необходим для метаболизма углеводов, белков и жиров, образования эритроцитов и благотворного влияния на функции нервной системы и печени.Около 80% витамина В6 содержится в мышцах, в печени, миокарде и в почках. Пиридоксин также принимает участие в поддержании натрий-калиевого баланса, уменьшает уровни гомоцистеина в крови, также необходим для синтеза нейротрансмиттеров серотонина, допамина, норадреналина и адреналина.
Дефицит пиридоксина сопровождается дефицитом магния и проявляется клиническими симптомами в виде раздражительности, заторможенности, снижении аппетита и тошноты, хейлоза, полиневритов, В6-зависимой тревожности [7].
Для диагностики гиповитаминоза наиболее часто прибегают к оценке концентрации пиридоксина в плазме крови. Величины в пределах 5–30 нг/мл считаются нормальными (коэффициент пересчета 4,05 (20–121 нмоль/л)), а при величинах менее 5 нг/мл х 4,05 (менее 20 нмоль/л) диагностируют дефицит. Однако, для достоверного установления дефицита пиридоксина измерение уровней его метаболитов в крови недостаточно.
Наиболее достоверным диагностическим тестом, отражающим обеспеченность любым витамином, является нагрузочный тест с использованием суточных доз. В ходе которого, определяется динамика концентраций витамина в плазме крови и в моче в ответ на прием его суточной дозы. При низкой обеспеченности пиридоксином наблюдается задержка витамина В6 в организме, о которой свидетельствуют неизменные уровни экскреции метаболитов витамина с мочой при нарастающих уровнях витамина В6 в плазме крови.
Несмотря на то, что пиридоксин встречается в большом количестве продуктов, дефицит пиридоксина широко распространен в России, особенно среди школьников [20]. Нормальная обеспеченность витамином В6 отмечается всего лишь у 36% детей (Рис. 7).
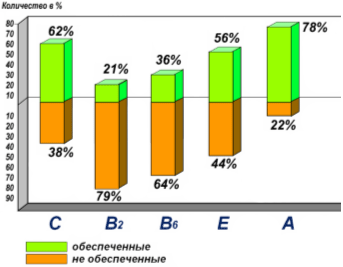
Рис. 7. Обеспеченность различными витаминами у московских школьников (по данным Спиричева В.Б., 2009).
Биологические эффекты пиридоксина обусловлены тем, что его производные являются кофакторами более 120 белков [7].
Экспериментальные исследования подтверждают тот факт, что дефицит пиридоксина, как и дефицит магния, ухудшает механические свойства соединительной ткани и приводит к дисплазии. У животных, отмечались черты гипотрофии соединительной ткани костей с эрозированной поверхностью [21]. В то же время, изучение эффектов анти-пиридоксиновых препаратов (4-деоксипиридоксин, пенициламин) показало, что эти ингибиторы биологических эффектов пиридоксина приводят к уменьшению стабильности коллагена [23] именно через ослабление связей между коллагеновыми волокнами, в т.ч. вследствие падения активности фермента лизилоксилазы [24].
В целом, экспериментальные исследования указывают на то, что пиридоксин явяляется эссенциальным микронутриентом для формирование здоровой соединительной ткани. Для установления точных молекулярных механизмов необходимо проведение систематического анализа данной проблемы с использованием современных биоинформационных технологий [25].
Заключение
Соединительная ткань является, в буквальном смысле, опорой всех тканей организма. Поэтому, недифференцированные дисплазии соединительной ткани у детей и подростков характеризуются широчайшим диапазоном проявлений, включающих детский и подростковый сколиоз, школьную миопию, пролапс митрального клапана, плоскостопие, варикозное расширение вен, нарушения фиксации органов (гастроптоз, нефроптоз, колоноптоз) и многие другие.
Основами реабилитации детей с патологией соединительной ткани являются кинезотерапия и диетология. В метаболической поддержке функции соединительной ткани большое значение имеют магний и пиридоксин. При дефиците как магния, так и пиридоксина, белковый синтез соединительной ткани замедляется. В случае дефицита магния, активность ММП увеличивается и внеклеточная матрица прогрессивно деградирует, так как структурная поддержка ткани (коллагеновые волокна, гелеобразная среда) разрушается быстрее, чем синтезируется. И наоборот, состояние соединительной ткани будет улучшатся, когда активности коллагеназ и эластаз, а также биосинтетических ферментов глюкозаминогликанов (гиалоуронансинтетаз, гиалоуронидаз, бета-галактозидаз) сбалансированы. Вследствие широкого распространения дефицитов и магния, и пиридоксина важным фактором в достижении такого баланса является достаточная обеспеченность магнием и пиридоксином.
Как видно из представленного выше анализа, магний оказывает разностороннее воздействие на метаболизм соединительной ткани. Образно, магний можно сравнить с дирижером соединительно-тканного биомолекулярного оркестра. От слаженности этого «оркестра» во все периоды детства и подросткового возраста зависит здоровье организма пациента во все последующие годы. Диагностика и лечение нДСТ с раннего детства, рациональное питание и культура движения могут полностью предотвратить развитие заболеваний соединительной ткани в зрелом возрасте или, по крайней мере, значительно облегчить их течение и улучшит качество жизни подрастающего поколения Россиян.
Литература
1. Баранов А.А. Сб. тр. XIV Конгресс педиатров России «Актуальные проблемы педиатрии», 15-18 февраля, 2010.
2. Нечаева Г.И., Яковлев В.М., Громова О.А., Торшин И.Ю. с соавт. Дисплазии соединительной ткани у детей и подростков. Инновационные стационар-сберегающие технологии диагностики и лечения в педиатрии. Союз Педиатров России, 2009, М., 96С.
3. Головской Б.В., Усольцева Л.В., Ховаева Я.В., Иванова Н.В.// Клин. Мед. – 2002. - № 80(12). - С. 39-41.
4. Яковлев В.М. Кардио-респираторные синдромы при дисплазии соединительной ткани / В.М. Яковлев, Г.И. Нечаева. - Омск. - 1994 - 217 с.
5. Громова О.А., Егорова Е.Ю., Торшин И.Ю., Федотова Л.Э., Юдина Н.В. Перспективы применения магния в педиатрии и детской неврологии. Журн.Педиатрия им. Сперанского, 2010, №7.
6. Школьникова М.А. Метаболизм магния и терапевтическое значение его препаратов. - М.: ИД Медпрактика – М. – 2002.
7. Torshin I.Yu., Gromova O.A. Magnesium and pyridoxine: fundamental studies and clinical practice. Nova Science, 2009, ISBN-10: 1-60741-704-9. 250pp.
8. Нечаева Г.И., Яковлев В.М., Конев В.П., Друк И.В., Морозов С.Л.. Дисплазия соединительной ткани: основные клинические синдромы, формулировка диагноза, лечение. Лечащий врач, 2010, № 5.
9. Викторова И.А.Методология курации пациентов с дисплазией соединительной ткани семейным врачом в аспекте профилактики ранней и внезапной смерти: Дисc. …док. мед. наук / И.А. Викторова – Омск, 2005. – 432 с.
10. Сohen М. М. The Сhild with Multiрle Birth Defeсts. 2nd Ed, New York, 1997.
11. Торшин ИЮ, Громова ОА. Молекулярные механизмы магния и дисплазии соединительной ткани. Росс. Мед. Ж., 2008, с. 263-269.
12. Alberts B, Johnson A, Lewis J, Raff M, Roberts R, Walter P. Molecular Biology of the Cell, 4th edition, Garland Science, 2002, ISBN 0815340729.
13. Okajima T, Fukumoto S, Furukawa K, Urano T. Molecular basis for the progeroid variant of Ehlers-Danlos syndrome. Identification and characterization of two mutations in galactosyltransferase I gene. J Biol Chem. 1999;274(41):28841-28844.
14. Di Ferrante N, Leachman RD, Angelini P, Donnelly PV, Francis G, Almazan A. Lysyl oxidase deficiency in Ehlers-Danlos syndrome type V. Connect Tissue Res. 1975;3(1):49-53.
15. Капелько В.И. Внеклеточный матрикс миокарда и его изменения при заболеваниях сердца //Кардиология. – 2000. – №9. – С. 78-90.
16. Mio K, Carrette O, Maibach HI, Stern R. Evidence that the serum inhibitor of hyaluronidase may be a member of the inter-alpha-inhibitor family. J Biol Chem. 2000;275(42):32413-32421.
17. Ueshima K, Shibata M, Suzuki T, Endo S, Hiramori K. Extracellular matrix disturbances in acute myocardial infarction: relation between disease severity and matrix metalloproteinase-1, and effects of magnesium pretreatment on reperfusion injury. Magnes Res. 2003;16(2):120-126.
18. Ahvazi B, Boeshans KM, Rastinejad F. The emerging structural understanding of transglutaminase 3. J Struct Biol. 2004;147(2):200-207.
19. Gacheru SN, Trackman PC, Shah MA, O'Gara CY, Spacciapoli P, Greenaway FT, Kagan HM. Structural and catalytic properties of copper in lysyl oxidase. J Biol Chem. 1990;265(31):19022-19027.
20. Спиричев В.Б. Витамины и интеллектуальное развитие детей. IV Российский форум «Здоровое питания с рождения: медицина, образование, пищевые технологии». СПб, 26-27 ноября, 2009.
21. Masse PG, Pritzker KP, Mendes MG, Boskey AL, Weiser H.Vitamin B6 deficiency experimentally-induced bone and joint disorder: microscopic, radiographic and biochemical evidence.Br J Nutr. 1994 Jun;71(6):919-32.PMID: 8031739
22. Masse PG, Yamauchi M, Mahuren JD, Coburn SP, Muniz OE, Howell DS.Connective tissue integrity is lost in vitamin B-6-deficient chicks.J Nutr. 1995 Jan;125(1):26-34.PMID: 7815173
23. Kajiwara T. Effect of D-penicillamine on vitamine B6 and collagen metabolism.Nippon Seikeigeka Gakkai Zasshi. 1979 Nov;53(11):1595-605. PMID: 528798
24. Bird TA, Levene CI.The effect of a vitamin B-6 antagonist, 4-deoxypyridoxine, on the cross-linking of collagen in the developing chick embryo.Biochem J. 1983 Mar 15;210(3):633-8.PMID: 6135414
25. Torshin I.Yu. Bioinformatics in the post-genomic era: sensing the change from molecular genetics to personalized medicine. Nova Biomedical Books, NY, USA, 2009, In “Bioinformatics in the Post-Genomic Era” series, ISBN: 978-1-60692-217-0.
Комментарии
ПРАКТИКА ПЕДИАТРА



