Фенотропил как рецепторный модулятор синаптической нейропередачи
СтатьиОпубликовано в журнале:
«Атмостфера. Нервные болезни», 2007, №4, с. 1-4
Г.И. Ковалев, В.И. Ахапкина, Д.А. Абаимов, Ю.Ю. Фирстова
Георгий Иванович Ковалев - докт. мед. наук, профессор, зав. лабораторией радиоизотопных методов исследований ГУ НИИ фармакологии им. В.В. Закусова РАМН.
Валентина Ивановна Ахапкина - директор департамента экспериментальной и клинической фармакологии, зам. генерального директора ЗАО “Отечественные лекарства”.
Денис Александрович Абаимов - канд. биол. наук, научный сотрудник лаборатории радиоизотопных методов исследований ГУ НИИ фармакологии им. В.В. Закусова РАМН.
Юлия Юрьевна Фирстова - аспирант лаборатории радиоизотопных методов исследований ГУ НИИ фармакологии им. В.В. Закусова РАМН.
Препарат Фенотропил (N-карбамоил-метил-4-фенил-2-пирролидон) по своим фармакологическим эффектам принадлежит к ноотропным лекарственным средствам. Большинство современных классификаций ноотропных средств относят производные пирролидона – рацетамы (пирацетам, оксирацетам, анирацетам и др.) – к так называемым “истинным ноотропам”, для которых улучшение мнестических функций – памяти, обучаемости – является главным (часто единственным) или наиболее выраженным проявлением психофармакологической активности. Эксперименты на лабораторных животных убедительно показывают, что рацетамы оказывают влияние на функционирование основных нейромедиаторных систем мозга – холинергическую, адренергическую, дофаминергическую, ГАМКергическую и глутаматергическую, причем в том направлении, “в котором эти системы имеют отношение к памяти и адаптации организма к экстремальным воздействиям” [1]. Тем не менее до последнего времени оставалось непонятным, через какие молекулярные мишени и синаптические механизмы рацетамы, включая Фенотропил, осуществляют эти действия. Более того, с момента опубликования в 1985 г. работы Bering and Muller [6] считалось общепринятым, что препараты этой группы не действуют ни на рецепторы нейромедиаторов, ни на какие-то другие места связывания, отличные от них [2, 12].
Однако целый ряд нейрохимических данных позволял сомневаться в справедливости данного утверждения. В частности, было экспериментально показано, что:
- пирацетам модулирует секрецию D-аспартата из изолированных нервных окончаний коры мозга крыс, вовлекая пресинаптические глутаматные рецепторы предположительно неNMDA-типа [4];
- анирацетам аллостерически потенцирует неNMDA-тип глутаматных рецепторов в ооцитах морского ежа [14];
- анирацетам увеличивает число рецепторов неNMDA-типа в мембранах коры мозга крыс [7];
- пирацетам модулирует число рецепторов глутамата в мембранах мозга крыс [3].
В последующие годы были открыты и подробно охарактеризованы новые фармакологические типы рецепторов нейромедиаторов, поэтому целью настоящей работы стало изучение влияния Фенотропила на различные типы рецепторов основных нейромедиаторов ЦНС.
Материал и методы исследования
Для количественного определения рецепторов в мембранных препаратах, приготовленных из компетентных морфофункциональных образований (гиппокампа, полосатого тела, коры больших полушарий) мозга крыс, применяли общепринятый метод радиорецепторного анализа с использованием специфических лигандов, меченных по тритию, в подборе которых основывались на рекомендациях IUPHAR от 2006 г. [5] (табл. 1).
Таблица 1. Типы рецепторов и использованные [3H]-лиганды
| Тип рецептора | Специфический лиганд, удельная активность (кюри/ммоль) |
| Дофаминовый, D1 | [G-3H]-SCH 23390, (60) |
| Дофаминовый, D2 | [G-3H]-спироперидол, (95) |
| Дофаминовый, D3 | [G-3H]-7-ОН-DРАТ, (120) |
| Серотониновый, HT2 | [G-3H]-кетансерин, (60) |
| Глутаматный, NMDA | [G-3H]-МК-801, (210) |
| Ацетилхолиновый, никотиновый | [G-3H]-(-)-никотин, (140) |
Были использованы два методических подхода в действии Фенотропила на рецепторы нейромедиаторов: метод in vitro и метод ex vivo. В первом случае мембраны мозга, содержащие рецепторы, инкубировали в пробирках вместе с радиоактивно меченным лигандом определенного рецептора в присутствии различных концентраций Фенотропила. Измерение специфически связавшегося в данных условиях радиоактивного лиганда с мембранным рецептором давало ответ на вопрос: оказывает ли лекарственное вещество прямое, конкурирующее действие на изучаемый тип рецептора? Активность препарата в данном варианте опытов выражали величиной концентрации, изменяющей специфическое связывание радиоактивного лиганда с рецептором на 50%, – IC50 (“inhibitory concentration 50”).
Во втором случае Фенотропил вводили крысам внутрибрюшинно в дозе 100 мг/кг, один раз в день в течение 7 сут. Группе контрольных животных вводили физиологический раствор. Через 24 ч после последней инъекции крыс декапитировали, извлекали мозг, готовили мембраны выделенных структур мозга, образцы которых инкубировали в присутствии радиолиганда. Обработанные результаты измерений были представлены величинами Bmax и Kd, отражающими концентрацию рецепторов в мембранах и степень структурного сродства (аффинитета) лекарственного средства к рецептору соответственно. Анализ данных ex vivo позволяет интерпретировать полученные сведения в свете возможного непрямого, модулирующего влияния Фенотропила на рецепторный профиль, отражающий реакцию клеток мозга на субхроническое введение препарата. Все меченые лиганды были получены методом твердофазного катализа в ОХВАВ Института молекулярной генетики РАН (заведующий – академик РАН Н.Ф. Мясоедов). Результаты экспериментов обрабатывали с помощью программ Statistica 6.0 и GraphPad 4.0 Prism.
Радиорецепторный анализ in vitro
Концентрационная зависимость прямого влияния Фенотропила на глутаматные рецепторы NMDA-типа представлена на рис. 1а. Видно, что влияние селективного антагониста этого типа рецептора МК-801 (дизоцилпина) описывается классической кривой (пунктирная линия) и характеризуется величиной IC50 0,007 мкмоль/л (табл. 2).
Рис. 1. Влияние Фенотропила на рецепторы in vitro. а – NMDA в гиппокампе, б – нАХ в коре.
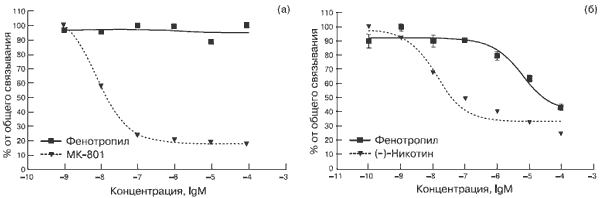
Таблица 2. Влияние Фенотропила на рецепторы нейромедиаторов in vitro
| Тип рецептора | Вещество сравнения, IC50 (мкмоль/л) | Фенотропил, IC50 (мкмоль/л) |
| Дофаминовый, D1 | SKF38393, 1,104 | >1000 |
| Дофаминовый, D2 | Галоперидол, 0,573 | >1000 |
| Дофаминовый, D3 | 7-OH-DPAT, 0,015 | >1000 |
| Серотониновый, HT2 | Кетансерин, 0,702 | >1000 |
| Глутаматный, NMDA | МК-801 (дизоцилпин), 0,007 | >100 |
| Ацетилхолиновый, никотиновый | (-)-Никотин 0,013 | 5,86 |
Внесение Фенотропила не обнаруживает эффекта конкуренции с лигандом за рецепторные места связывания вплоть до максимальной использованной концентрации 100 мкМ (сплошная кривая), что интерпретируется как отсутствие структурного сродства препарата к глутаматному рецептору NMDA-типа.
Аналогичная инертность Фенотропила продемонстрирована в отношении всех трех типов дофаминовых рецепторов и рецептора серотонина 5-HT2-типа: в пределах использованных концентраций препарат не вмешивался в процесс связывания специфических рецепторных лигандов SKF38393 (D1-тип), галоперидола (D2-тип), 7-OH-DPAT (D3-тип) и кетансерина (5-HT2-тип) вплоть до 1 ммоль/л (см. табл. 2).
Напротив, изучение влияния Фенотропила на рецепторное связывание высокоизбирательного (IC50 13 нмоль/л) лиганда никотинового типа холинорецепторов [G-3H]-(-)-никотина позволило обнаружить фармакологически значимую конкуренцию препарата за данный тип рецептора с IC50 5,86 мкмоль/л (см. рис. 1б и табл. 2).
Таким образом, представитель структурной группы производных пирролидона Фенотропил является н-холинергическим препаратом прямого рецепторного действия.
Радиорецепторный анализ ex vivo
Субхроническое введение Фенотропила в ноотропной дозе 100 мг/кг и последующий радиорецепторный анализ результатов связывания лигандов показали широкий спектр действия препарата. Графики насыщения рецепторов изученных типов в контрольной (пунктирная кривая) и опытной (сплошная кривая) группах крыс, а также рассчитанные величины Kd и Bmax представлены на рис. 2 и в табл. 3 соответственно.
Рис. 2. Влияние Фенотропила на рецепторы ex vivo. а - NMDA в гиппокампе, б - нАХ в коре, в - D1 в стриатуме, г - D2 в стриатуме, д - D3 в стриатуме, е - 5-HT2 во фронтальной коре.
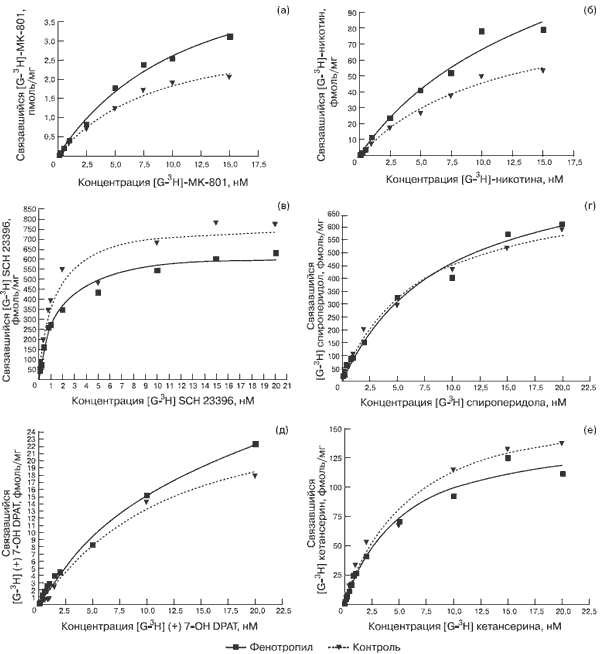
Таблица 3. Влияние Фенотропила на показатели рецепторного связывания в мозге крыс ex vivo после 7-кратного введения (m ± S.E.)
| Тип рецептора | Контроль (физраствор) | Фенотропил, 100 мг/кг/сут | ||
| Kd, нМ | Bmax, фмоль/мг | Kd, нМ | Bmax, фмоль/мг | |
| D1 | 1,14 ± 0,26 | 778,8 ± 45,3 | 1,45 ± 0,20 | 639,8 ± 23,2* |
| D2 | 6,87 ± 1,00 | 761,5 ± 44,0 | 10,67 ± 2,66 | 931,5 ± 102,7 |
| D3 | 14,70 ± 3,44 | 32,1 ± 4,2 | 17,31 ± 2,57 | 41,5 ± 3,5* |
| HT2 | 5,65 ± 0,94 | 176,6 ± 11,0 | 4,98 ± 0,83 | 148,3 ± 8,9* |
| NMDA | 8,3 ± 1,5 | 3350 ± 300 | 11,2 ± 2,0 | 5540 ± 530* |
| нAX | 13,1 ± 3,2 | 103,3 ± 14,5 | 13,8 ± 4,9 | 161,1 ± 33,5* |
* Различия с показателями в контроле (р
Продемонстрировано, что в ответ на 7-кратное системное введение Фенотропила мозг опытной группы животных индуцирует статистически значимые изменения для большинства из исследованных типов рецепторов (см. рис. 2а–2в, 2д, 2е).
Важно отметить, что ни для одного из них изменения не касаются величины Kd, отражающей степень сродства рецептора к специфическому лиганду, тогда как концентрация рецепторов (Bmax) меняется как в сторону увеличения, так и в сторону уменьшения (см. табл. 3).
Так, под влиянием Фенотропила более всего увеличилась концентрация NMDA-рецепторов в гиппокампе (+65% относительно группы плацебо, см. рис. 2а), никотиновых рецепторов в коре мозга (+56%, см. рис. 2б) и D3-дофаминовых рецепторов в стриатуме (+29%, см. рис. 2д). Относительное уменьшение количества рецепторов произошло в стриатуме для D1-дофаминовых рецепторов (–18% в сравнении с группой плацебо, см. рис. 2в) и в коре мозга для серотониновых рецепторов типа HT2 (–16%, см. рис. 2е). Отсутствие заметных отличий в количестве D2-рецепторов можно объяснить тем, что использованный в качестве D2-лиганда спироперидол также проявляет некоторое сродство к субпопуляциям D3- и HT2-рецепторов, в связи с чем суммарное связывание не выявило отличий.
Следовательно, субхроническое введение Фенотропила приводит к разнонаправленным изменениям в количестве нейромедиаторных рецепторов по сравнению с плацебо: количество никотиновых, NMDA и D3-дофа-миновых типов рецепторов увеличивается, а количество HT2-серотониновых и D1-дофаминовых – уменьшается.
Заключение
Представленные в данной работе данные о рецепторном механизме действия Фенотропила, с одной стороны, хорошо согласуются со сформировавшимися к сегодняшнему дню взглядами об основных нейрохимических мишенях ноотропных средств рацетамового ряда, с другой стороны, привносят в них абсолютно новые элементы. Так, прямое или опосредованное вовлечение дофаминовых рецепторов в механизмы когнитивной деятельности и эффекты ноотропных препаратов обсуждается в работах авторов последних лет [11], равно как и роль в этих процессах серотониновых рецепторов HT2-типа [13].
Изученный препарат Фенотропил оказался активным в первую очередь в отношении рецепторов никотинового и NMDA-типа глутаматных рецепторов. Но если в отношении первых препарат обнаружил прямое действие, то увеличение концентрации вторых можно объяснить только через механизм нейромодуляции. Ряд свидетельств в пользу участия глутаматных рецепторов NMDA- и неNMDA-типов в процессах памяти и обучения, а также в механизме действия рацетамов приведен в начале данной статьи.
Холинергические механизмы пластичности нервных клеток являются признанной мишенью для ноотропных препаратов, в основе чего лежат данные нейрофизиологии, нейрохимии, нейрофармакологии и клиники. Во-первых, никотиновые рецепторы активно участвуют в когнитивных процессах [17, 18]. Во-вторых, н-холинергические элементы вовлечены в патогенетические механизмы расстройств памяти [8, 10, 15, 16]. В-третьих, одной из основных групп препаратов антиамнестического действия являются средства, воздействующие на никотиновые рецепторы. Средства, блокирующие активность ацетилхолинэстеразы (амиридин, такрин), и холинергические предшественники (глиатилин) активно используются в клинике против болезни Альцгеймера [9]. Дальнейшее уточнение рецепторных механизмов действия Фенотропила может быть получено при изучении аналогичного рецепторного профиля при моделировании нарушений когнитивной деятельности.
Список литературы
1. Ахапкина В.И., Воронина Т.А. // Фарматека. 2005. Т. 13. С. 1.
2. Ковалев Г.В. Ноотропные средства. Волгоград, 1990.
3. Ковалев Г.И. Изучение роли межмедиаторных взаимодействий в механизме формирования эффектов ноотропных средств: Автореф. дис. … докт. мед. наук. М.,1993.
4. Прихожан А.В., Ковалев Г.И. // БЭБиМ. 1986. № 10. С. 440.
5. Аlexander S.P.H. et al. // B r. J. Pharmacol. 2006. V. 147. Suppl. 3. P. S1. ..
6. Bering B., Muller W.E. // Arzneimittel. Forsch. 1985. V. 35. № 9. P. 1350.
7. Copani et al. // J. Neurochem. 1992. V. 58. P. 1199.
8. Dwoskin L.P., Crooks P.A. // JPET. 2001. V. 298. P. 395.
9. Ellis J.M. // JAOA. 2005. V. 105. № 3. P. 145.
10. Francis P. T. et al. // J. Neurol. Neurosurg. Psychiatry. 1999. V. 66. P. 13.
11. Glickstein S. et al. // J. Neurosci. 2002. V. 22. № 13. P. 5619.
12. Gouliaev A., Senning A. // Brain. Res. Rev. 1994. V. 19. № 2. P. 180.
13. Harvey J.A. // Learn. Mem. 2003. V. 10. P. 355.
14. Ito I. et al. // J. Neurochem. 1990. V. 424. P. 533.
15. Mesulam M. // Learn. Mem. 2004. V. 11. P. 43.
16. Parent M.B., Baxter M.G. // Learn. Mem. 2004. V. 11. P. 9.
17. Pepeu J., Giovannini M.L. // Learn. Mem. 2004. V. 11. P. 21.
18. Tinsley M.R. et al. // Learn. Mem. 2004. V. 11. P. 3.
Комментарии
ПРАКТИКА ПЕДИАТРА



