Анализ молекулярных механизмов воздействия железа (ii), меди, марганца в патогенезе железодефицитной анемии
Статьи Опубликовано в журнале:«Клиническая фармакология и фармакоэкономика» №1, 2010, Москва
О.А. Громова1, 2, И.Ю. Торшин1, А.К. Хаджидис3
1Российский Сотрудничающий Центр Института Микроэлементов ЮНЕСКО, Москва, 2 ГОУ ВПО Ивановская государственная медицинская академия МЗРФ, 3 ГОУ ВПО Санкт-Петербургская государственная педиатрическая академия МЗРФ
Железодефицитная анемия (ЖДА), несмотря на свое название, не является результатом только дефицита железа. Любой микроэлемент, в том числе и железо, оказывает свои биологические функции в контексте многих других микроэлементов. Существует целый спектр микронутриентов, необходимых для полноценного усвоения железа организмом и проявления биологических эффектов железа. В настоящем исследовании проведен анализ патофизиологии ЖДА с точки зрения потребности в различных микронутриентах. Детализируется синергизм между железом, марганцем и медью на уровне молекулярных маршрутов и обсуждаются фармакологические особенности различных форм железа. Результаты дают обоснование для эффективного патогенетического лечения ЖДА и профилактики дефицита железа во время беременности.
Ключевые слова: патофизиология железодефицитной анемии, медь, марганец, молекулярные механизмы
Key words: pathophysiology iron deficiency anemia, copper, manganese, molecular mechanisms
Введение
Дефициты железа и йода наиболее широко распространены. Принято считать, что эти дефициты являются монодефицитами, т. е. обусловлены недостатком в основном только одного микроэлемента. Однако, в организме гомеостаз того или иного микроэлемента не существует в отрыве от гомеостаза многих других макро- и микроэлементов [1]. Например, невозможно добиться устойчивых результатов лечения йод-дефицита назначением только препаратов йода; селен, медь и железо необходимы для наиболее эффективной и безопасной терапии [2].
Детальные молекулярные механизмы биологического действия микроэлементов остаются вне внимания большинства исследователей [3]. В частности это касается этиологии и терапии железодефицитной анемии (ЖДА) и анемии беременных (АБ).
Упрощенный взгляд на этиологию и терапию ЖДА пренебрегает фактом, что на гомеостаз железа влияют более 200 белков и соответствующих генов, кодирующих эти белки [4]. При наличии у пациента определенных вариантов генов (генетических полиморфизмов или редких мутаций), гомеостаз железа значительно затрудняется. Поэтому у таких пациентов может быть повышенный риск возникновения ЖДА даже при нормальном потреблении железа.
Эти 200 и более белков гомеостаза железа не могут функционировать без соответствующих кофакторов (таких, как производные витаминов группы В, ионов меди, марганца, цинка и др.). Поэтому дефицит этих кофакторов, подобно генетическим факторам, также может способствовать функциональному дефициту железа.
Фармакокинетика и фармакодинамика усвоения железа изменяется в зависимости от дополнительных факторов. Например, совместный прием препаратов железа и цинка в растворе ухудшает усвоение и железа, и цинка [4]. Присутствие в пище фитатов, воспаление, дефицит меди и избыток кальция снижают усвоение железа [2].
ЖДА является комплексным патофизиологическим процессом и приводит к недостаточному поступлению кислорода к тканям. Железо необходимо для синтеза гема, входящего в состав гемоглобина — белка-переносчика кислорода. Помимо железа, в биохимических каскадах синтеза гема участвуют многие кофакторы (производные витаминов В6, РР и В5). Гемоглобин активен в составе эритроцитов, так что на анемию влияют интенсивность клеточных процессов гемопоэза и гемолиза. Медь и марганец — интегральные составляющие активных центров ферментов так называемой дыхательной цепи, т. е. ферментов, непосредственно вовлеченных в усвоение молекулярного кислорода. Медь и марганец также являются кофакторами ключевых белков гомеостаза железа, и гомеостаз железа сильно «обкрадывается» при дефиците этих микроэлементов. Иначе говоря, железо — безусловно важный, но не единственный фактор, определяющий возникновение и течение желе-зодефицитной анемии.
Еще в прошлом века стало известно, что для максимально эффективной терапии ЖДА необходимы не только железо, но и медь, марганец и аминокислоты. Много лет проводились фармакологические исследования по подбору наиболее эффектных пропорций Fe:Cu:Mn, например, в работе 1935—1936 гг. было проанализировано роль соотношения Fe:Cu при восстановлении гемоглобина при «пищевой анемии» [6]. Однако фундаментальные молекулярные механизмы, через которые осуществляется воздействие этих микроэлементов на патогенез ЖДА, не были исследованы.
Микронутриенты, поддерживающие функции железа на молекулярном уровне
Используемый в данной работе метод анализа функциональных взаимосвязей [7] основан на постоянно обновляющейся аннотации генома и позволяет проводить систематические анализы взаимодействий различных биологических лигандов (будь-то нутриенты, микронутриенты, лекарства, ксенобиотики и др.) друг с другом и с белками протеома человека. Разработанное нами программное обеспечение позволило проанализировать аннотацию генома и все имеющиеся электронные библиотеки белков. Метод позволил установить спектр микронутриентов, непосредственно необходимых для поддержания биологических эффектов железа и сформулировать соответствующие молекулярные механизмы, с указанием конкретных белков и генов (табл. 1).
Таблица 1. Микронутриенты необходимые для поддержания биологических функций железа
| Нутриент | Число белков |
|
Витамины и их производные |
|
| Флавинадениндинуклеотид (ФАД, производное витамина В2) | 17 |
| Аскорбиновая кислота (витамин С) | 12 |
| Флавинмононуклеотид (ФМН, производное витамина В2) | 5 |
| Пиридоксаль фосфат (производное витамина В6) | 5 |
| Микроэлементы | |
| Mn 2+ | 22 |
| Zn 2+ | 21 |
| Cu+/Cu2+ | 9 |
| MoO2-4 | 3 |
В дальнейшем обсуждаются взаимосвязи микроэлементов с метаболизмом железа. Особое внимание уделяется взаимодействиям марганца и меди, так как это имеет особое значение для фармакотерапии анемических состояний.
Марганец — эссенциальный микроэлемент и кофактор более 200 белков. В клетках значительная часть запасов марганца сосредоточена в митохондриях, поэтому марганцем наиболее богаты печень, почки, плацента, поджелудочная железа, кишечник. Пищевыми источниками марганца являются чай, стручковые, орехи, клюква, черника. Марганец всасывается в кишечнике, всасыванию препятствуют соединения кальция, избыток железа, фосфаты, оксалаты. При дефиците железа, наоборот, биоусвояемость марганца повышается [2].
Адекватный уровень потребления марганца: 2 мг/сут. При недостаточном потреблении марганца и нарушениях гомеостаза этого элемента возникает дефицит марганца (рубрикация Е61.3 по МКБ-10).
Анализ функциональных взаимосвязей между функциями железа и марганца показал, что марганец влияет на функцию 22 белков, вовлеченных в гомеостаз железа. Эти белки можно сгруппировать в 5 биохимических функциональных групп. Каждый из этих белков связывает либо ион железа, либо гем, либо молекулярный кислород. Большинство белков вовлечены в ответ организма на состоянии гипоксии. Информация о белках и соответствующих генах суммирована в табл. 2.
Таблица 2. Функциональные группы Mn-зависимых белков, влияющих на биологические функции железа
| Биохимическая функциональная группа | Число белков | Биологическое значение |
| Серин-треонин фосфатазы | 14 | Внутриклеточная передача сигнала |
| Гуанилат-циклазы | 4 | Передача сигнала от окиси азота (NO) |
| Ион-транспортеры | 2 | Распределение Fe в печени и организме |
| Малат дегидрогеназа | 1 | Метаболизм сахаров |
| Митохондральная пептидаза | 1 | Импорт белков гомеостаза Fe в митохондрии |
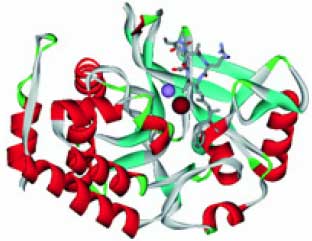
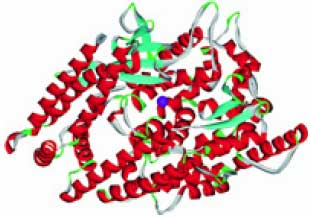
Серин-треонин фосфатазы (S/T-фосфатазы, гены PPP1CA, PPP2CA, PPP2CB, PPP4C, PPP5C, PPP6C и т. д.) ферменты, гидролизующие фосфорилированные формы остатков серина и треонина белков. Так как фосфорилирование и дефосфорилирование аминокислотных остатков повсеместно используется в клетке как способ передачи сигнала, фосфатазы модулируют передачу самых различных сигналов, имеющих принципиальное значение для выживания клетки. Например, активность S/T-фосфатазы 1 необходима для деления клетки, метаболизма гликогена, мышечного сокращения и синтеза белка. Так как и марганец, и железо необходимы для активности S/T-фосфатаз, то дефицит этих элементов будет приводить к замедлению роста клеток, в т. ч. эритроцитов. Модель пространственной структуры S/T-фосфатаз показана на рис. 1 (см. обл. стр. 3).
Рис. 1. Пространственная структура S/T-фосфатаз (S/T-фосфатазы 1). Показаны атом марганца (фиолетовая сфера), атом железа (темнокрасная сфера) и молекула ингибитора (решетчатая модель) в активном центре фермента (модель на основе PDB 3E7A, 3E7B)
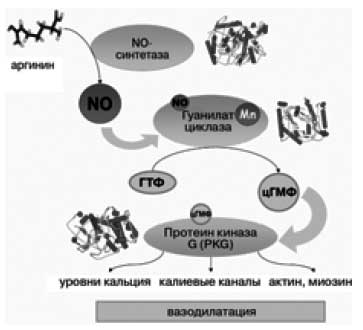
Гуанилат-циклазы (гены GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3) модулируют передачу сигнала от окиси азота (NO). NO, наименьшая из сигнальных молекул, диффундирует внутрь клеток гладкой мускулатуры и активирует гуанилат циклазу. Гуанилат циклаза производит циклический гуанозинмонофосфат (цГМФ) из гуанозинтрифосфата (ГТФ). Молекулы цГМФ активируют протеинкиназу G (PKG), фосфорилирующую белки, которые регулируют уровни кальция, чувствительность мышечной клетки к кальцию, гиперполяризацию клетки через калиевые каналы, актин, миозин, что и приводит, в конечном счете, к вазодилатации [8]. Менее известен факт, что гуанилат цикалазы также могут выступать как сенсор кислорода, тем самым непосредственно имея отношение к анемическим состояниям [9]. Для каталитической активности гуанилатциклаз необходим гем как кофактор; ионы марганца активируют фермент [10]. Пространственная структура полной молекулы гуанилат циклаз все еще не установлена.
Рис. 2. Молекулярные механизмы NO-опосредованной вазодилатации
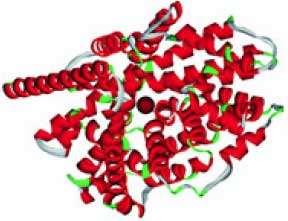
Ион-транспортеры типа DMT (транспортер двухвалентных металлов, гены SLC11A1, SLC11A2) важны для транспорта двухвалентных ионов металлов и прежде всего Fe и Mn. Эти ионные транспортеры вовлечены в адсорбцию железа в дуоденальных энтероцитах и также в транспорт железа внутрь клеток-предшественников эритроцитов. DMT-транспортеры влияют на накопление железа и марганца в печени и других органах. Дефекты в генах транспортеров приводят к гипохромной анемии (код по OMIM 206100), характеризующейся сниженным содержанием гемоглобина в эритроцитах и малым размером последних. Модель пространственной структуры транспортеров показана на рис. 3 (см. обл. стр. 3).
Рис. 3. Пространственная структура молекулы DMT-транспортеров (модель на основе PDB 2a65). Ион железа (темно красная сфера) в канале транспортера
Малат дегидрогеназа (ген MЕ3) — один из ферментов, вовлеченных в аэробное клеточное дыхание. Этот фермент расположен в матриксе митохондрии и связывает две ветви метаболизма сахаров: гликолиз и цикл трикарбоновых кислот (т. н. «цикл Кребса»). Малат дегидрогеназа имеет НАДФ (никотинамид динуклеотид, производное витамина РР) и ионы марганца в качестве кофакторов и катализирует превращение малата в пируват — основой субстрат цикла трикарбоновых кислот. Уровни активности различных форм малат-дегидрогеназ могут уменьшаться при железодефицитной анемии [11]. Пространственная структура фермента представлена на рис. 4 (см. обл. стр. 3).
Рис. 4. Пространственная структура активного центра субъединицы малат дегидрогеназы (PDB 1PJ3). Ион марганца (фиолетовая сфера), молекула пирувата и молекула НАДФ (решетчатая модель слева от марганца)
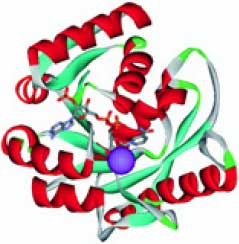
Митохондральная пептидаза (ген MIPEP) регулирует импорт различных белков в митохондрии, в т. ч. белков гомеостаза железа таких, как феррохелатаза и др. Функцией этой пептидазы является протеолиз N-концевых октапептидов импортируемых белков, что приводит к приобретению этими белками полной биологической активности (так называемое созревание полипептидов). Этот фермент является регулятором уровней железа: инициируя созревание транспортных белков железа, стимулирует накопление железа в митохондриях. При избытке железа активность фермента падает; избыточное падение активности фермента приводит к потере митохондриями железа [12]. Активность фермента стимулируется ионами марганца, магния или кальция. Модель пространственной структуры фермента показана на рис. 5 (см. обл. стр. 3).
Рис. 5. Пространственная структура митохондриальной пептидазы MIPEP (модель на основе PDB 2O36). Вероятное расположение иона марганца (фиолетовая сфера) в активном центре фермента
Медь — эссенциальный микроэлемент, имеющий прямое отношение к процессам дыхания: один из ключевых ферментов «дыхательной цепи» переноса электронов, цитохром С оксидаза, содержит ион меди и гем в качестве кофакторов. Поэтому можно считать медь одним из основных физиологических синергистов железа. В организме человека содержится 70—100 мг меди, 30% от этого количества — в печени. Хорошими пищевыми источниками меди являются мясо и субпродукты, стручковые, зерновые, орехи, зеленые овощи. Молибден увеличивает потерю меди с мочой. Цинк в ионной форме может конкурировать с медью за всасывание. В эксперименте дефицит меди непосредственно приводил к железодефицитной анемии вследствие нарушения абсорбции железа [13].
Норма потребления меди — 0,2—1,5 мг/сут, в зависимости от возраста. При нормальном питании дефицит меди встречается редко и возникает лишь у людей с белковой недостаточностью при голодании или при энтеритах с синдромом мальабсорбции. Дефицит меди (диагнозы по МКБ-10 Е 61.0 «недостаточность меди», Е 83.0 «нарушение обмена меди») приводит к развитию анемии, демиелинизации нервных клеток и потере пигментации кожи и волос, нарушению формирования сердечно-сосудистой системы, скелета у плода.
Медь принимает участие в азотном обмене, входя в состав нитрат-редуктазного комплекса, участвует в процессах, которые обеспечивают ткани кислородом. Медь входит в состав десятков ферментов, задействованных в окислительно-восстановительных реакциях, поддержке состояния соединительной ткани, синтезе нейротрансмиттеров, активации пептидных гормонов, каскаде свертывания крови, всасывании железа и др. К наиболее известным медьзависимым ферментам относятся Cu/Zn-супероксид дисмутаза (антиоксидантный эффект), цитохром С оксидаза (перенос электронов в дыхательной цепи митохондрий), лизил оксидаза (модификация коллагена, соединительная ткань), дофамин-β-гидроксилаза (синтез катехоламинов), тирозиназа (синтез меланина, пигментация и защитные свойства кожи) и др.
Анализ генома человека указал на существование 9 медьзависимых белков, которые относятся к четырем функциональным группам: цитохром С оксидаза, супероксид дисмутазы, металлоредуктазы STEAP и гефестин (табл. 3).
Таблица 3. Функциональные группы медь-зависимых белков, влияющих на биологические функции железа
| Биохимическая функциональная группа | Число белков | Биологическое значение |
| Металлоредуктазы STEAP | 4 | Абсорбция Fe посредством трансферрина |
| Цитохром с оксидаза | 2 | Фермент дыхательной цепи |
| Супероксид дисмутазы | 2 | Антиоксидантная защита, гомеостаз железа |
| Гефестин | 1 | Взаимопревращение Fe2+/Fe3+ |
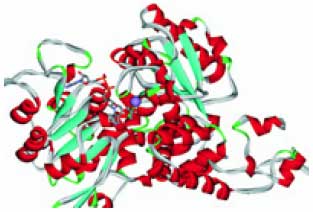
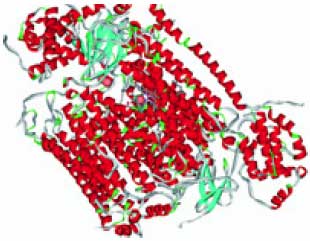
Металлоредуктазы типа STEAP (гены STEAP1, STEAP2, STEAP3, STEAP4), впервые обнаруженные как трансмембранные эпителиальные антигены простаты (отсюда и сокращение STEAP), являются эндосомальными ферриредуктазами, которые необходимы для эффективной абсорбции железа посредством трансферрина. Эти металлоредуктазы восстанавливают железо Fe3+ в биологически полезное двухвалентное Fe2+ и регулируют поглощение железа и меди клетками-предшественниками эритроцитов. Металлоредуктазы также восстанавливают двухвалентную медь Cu2+ в одновалентную Cu1+, являющуюся сильным восстановителем [14]. Недостаток активности металлоредуктазы STEATP4 связан с резистентностью к инсулину [15] и артритом через регулировку провоспалительных цитокинов [16]. Кофакторами ферментов являются ФАД и НАД производные витаминов В2 и РР соответственно. Модель пространственной структуры металлоредуктаз показана на рис. 6 (см. обл. стр. 3).
Рис. 6. Пространственная структура металлоредуктаз STEATP (модель на основе PDB 1jay). Кофактор (НАД) и предположи тельное расположение иона меди (сфера)
Цитохром С оксидаза (гены MTCO1, MTCO2) компонент дыхательной цепи, катализирующей восстановление молекулярного кислорода до воды. В ходе этого процесса перенос электронов и протонов способствует синтезу АТФ в митохондриях. Электроны переносятся через гем и атом меди, являющиеся кофакторами цитохром С оксидазы. Падение активности цитохром С оксидазы (вследствие генетических дефектов или глубокого дефицита железа и меди) приводит к наследственной нейропатии Лебера (номер 535000 по OMIM), идиопатической сидеробластической анемии (OMIM 516030) [17] и миоглобинурии (OMIM 550500). Пространственная структура фермента показана на рис. 7 (см. обл. стр. 3).
Рис. 7. Модель пространственной структуры цитохром с оксидазы (на основе PDB 1v54). Атомы меди (розоватые сферы) и гем (решетчатая модель) в активном центре фермента.
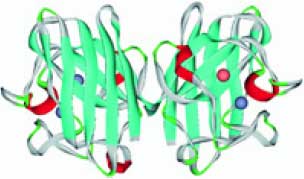
Cu,Zn-супероксид дисмутазы (гены SOD1, SOD2) ферменты антиоксидантной защиты организма. Деградируя пероксидный и надпероксидный анионы, супероксид дисмутазы, наряду с каталазой, обеспечивают защиту организма от высокореактивных форм кислорода. Удаляя свободные радикалы, супероксид дисмутазы способствует нормализации процессов воспаления. Наоборот, дефицит активности дисмутаз вследствие генетических дефектов или дефицит цинка и меди приводит к амиотрофическому латеральному склерозу нейродегенеративному заболеванию моторных нейронов (MIM 105400). Помимо влияния на процессы воспаления, активность супероксид дисмутаз также влияет на гемолиз, анемию и клеточный гомеостаз железа [18]. Пространственная структура супероксид дисмутаз показана на рис. 8 (см. обл. стр. 3).
Рис. 8. Пространственная структура димера (комплекса двух субъединиц) Cu,Zn-супероксид дисмутазы 1. Ион цинка (серо-синяя сфера) и ион меди (сфера розового цвета) в активном центре фермента (PDB 2VR8)
Гефестин - медьзависимый белок гомеостаза железа. Гефестин является ферроксидазой и обеспечивает взаимопревращения двух- и трехвалентного железа. Белок содержит 6 ионов меди и регулирует поток железа, связанный с транспортом через ферропортин. Модель пространственной структуры показана на рис. 9 (см. обл. стр. 3).
Рис. 9. Модель пространственной структуры гефестина (на основе PDB 1kcw). Возможные расположения ионов меди в структуре фермента (розовые сферы)
Фармакотерапия ЖДА
Патогенетической основой терапией железо-дефицитной анемии являются препараты железа. Однако, как показывают приведенные выше результаты анализа, биологические функции железа могут тормозиться в отсутствии целого ряда микронутриентов (особенно Zn, Mn, Cu, Mo, Cr, I, витаминов С, В2, В6, Вс, В12), причем следует отметить особую роль меди и марганца.
Можно предположить синергизм, т. е. повышение эффективности при совместном применении железа и указанных микронутриентов. В то же время такие витамины, как В2, В6, С, необходимые для поддержания функций железа, присутствуют в большинстве витаминно-минеральных комплексов (ВМК), причем в достаточных количествах.
Несмотря на синергизм между микроэлементами на уровне физиологических систем и конкретных белков, существует также определенный фармакокинетический антагонизм между железом, цинком, медью, молибденом и марганцем. Марганец всасывается в кишечнике, всасыванию препятствуют соединения кальция, избыток железа, фосфаты и оксалаты. При дефиците железа, наоборот, биоусвояемость марганца повышается. Железо, медь, магний ухудшают всасывание цинка. Молибден увеличивает потерю меди с мочой, а цинк может конкурировать с медью за всасывание. Из этих фармакокинетических антагонизмов ясно, что наиболее приемлем совместный прием железа, меди и марганца (фармакокинетический антагонизм минимален), а цинк и молибден следует принимать отдельно от железа, меди и марганца.
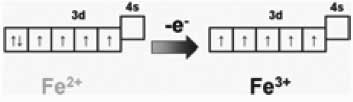
При выборе препарата железа следует помнить о различии между трех- и двухвалентным железом: в растворе двухвалентное железо — восстановитель, трехвалентное железо — окислитель [19]. Двухвалентное железо (Fe2+) может отдать один электрон и превратится в трехвалентное (Fe3+). При нормальных условиях, +3 — высшая степень окисления железа в растворе, так что в организме трехвалентное железо никогда не может быть восстановителем. В то же время трехвалентное железо является акцептором протонов (т. е. окислителем) и восстанавливается до двухвалентного: Fe3++ е– = Fe2+ [20]. Это процесс можно проиллюстрировать с использованием схем распределения электронов по энергетическим уровням (рис. 10).
Рис. 10. Окисление двухвалентного железа в трехвалентное
К сожалению, многие авторы [21] предоставляют достаточно подробный анализ железодефицитной анемии, включая этиологию и диагностику заболевания, и заявляют, что двухвалентное железо восстанавливается в трехвалентное. Цитируем дословно: «при своем восстановлении в трехвалентное железо в слизистой оболочке ЖКТ двухвалентные соли железа образуют свободные радикалы». Не верно и заявление авторов о том, что гидроксиды и оксиды есть наилучшие фармакологические формы железа.
Следует помнить, что при своем восстановлении в двухвалентное железо в слизистой оболочке ЖКТ трехвалентные соли железа образуют свободные радикалы, что может привести к развитию оксидативного стресса и, в частности, вызвать побочные реакции со стороны органов пищеварения (диспепсические явления, металлический привкус во рту, потемнение зубов и десен, тошнота, рвота, чувство переполнения желудка, запор, диарея). Исследования клеток в культуре показали, что уровни антиоксидантных ферментов супероксид дисмутаз и глутатиона были гораздо выше в клетках, обработанных двухвалентным, а не трехвалентным железом [22]. Кроме того, двухвалентное железо всасывалось более эффективно, чем трехвалентное (р Следует помнить о наиболее приемлемых фармацевтических формах железа и других микроэлементов. В большинстве биологически активных добавок и в минерализованных продуктах питания используется сульфат железа, который отличается низкой биоусвояемостью и способствует повреждениям сосудов и других тканей. Более того, сульфат железа вызывает желудочно-кишечные расстройства и врожденные аномалии у плода. Поэтому ВОЗ рекомендует беременным препараты на основе фумарата железа либо других безопасных органических солей железа [24]. Добавки с органическим железом глюконатом железа, фумаратом железа, цитратом железа не нейтрализуют витамин Е. Они представлены в различных дозировках, обычно до 320 мг. Эти формы не вызывают запора и легко переносятся при повышенной чувствительности. В настоящее время прослеживаются отчетливая тенденция удаления железа из витаминно-минеральных комплексов (особенно для пожилых пациентов) и применение специальных препаратов железа.
Препараты железа перорального применения более предпочтительны по сравнению с медикаментами парентерального введения. Терапевтический эффект от препаратов железа, назначаемых физиологически, т. е. реr os, наступает несколько позже, чем при назначении внутривенно или внутримышечно, но побочные эффекты при назначении внутрь возникают существенно реже.
Дозировка препарата имеет большое значение для эффективности и безопасности терапии. В рекомендациях ВОЗ указано, что профилактическое применение препаратов железа (60 мг/сут) необходимо всем беременным. Его необходимо начинать как можно раньше (не позднее третьего месяца) и продолжать до конца беременности. Если у женщины уже имеется анемия, то препараты железа целесообразно применять в лечебной дозе (120 мг). Однако, увеличение суточной дозы свыше 200 мг (в пересчете на элементарное железо) значительно повышает частоту и выраженность побочных реакций.
Клинические исследования сочетанных препаратов железа
Факты, касающиеся биологического синергизма между железом и определенными микронутри-ентами, представлены выше. Исследования, в которых проводилась терапия органическими препаратами двухвалентного железа и сочетанная терапия железом и другими микронутриентами (например, медью и марганцем [25], В12 и микроэлементами [26] и др.), показали высокую эффективность и безопасность органических солей и успех сочетанной терапии.
Исследование эффективности и переносимости питьевой формы глюконата железа (II) у 40 беременных, страдающих железодефицитной анемией (гемоглобин Препарат «Тотема» — сочетанная форма глюконата железа с глюконатом меди и глюконатом марганца. Применение препарата при предоперационной подготовке у 86 женщин с железодефицитной анемией (гемоглобин 88—100 г/л, уровень сывороточного железа — 8—12 мкмоль/л, цветовой показатель — 0,8—0,9), страдающих миомой матки, показало эффективность терапии при полном отсутствии побочных эффектов. Даже при сверхкоротком курсе (5 до 10 дней) уровень гемоглобина повышался на 12—15 г/л, уровень сывороточного железа — на 4—8 мкмоль/л. Короткие курсы приемы сделали показатели крови достаточными для производства оперативного лечения и, тем самым, устранения причины анемии [28].
Исследование эффективности и возможных побочных эффектов препарата «Тотема», назначаемого с целью коррекции железодефицитных состояний, у 20 беременных показало достоверное повышение уровня гемоглобина (р Заключение
Результаты проведенного в настоящей работе анализа физиологических синергизмов железа показывают, что по крайней мере 10 других микронутриентов необходимы для более эффективного проявления биологических эффектов железа: марганец, медь, молибден, хром, йод и витамины С, В2, В6. Результаты анализа позволяют предположить, что совместное назначение органических солей двухвалентного железа в сочетании с солями марганца и меди будет обладать минимумом фармакологического антагонизма и максимумом адсорбции в организме. В отличие от прооксидантного трехвалентного железа (и в особенности неорганических форм, таких, как сульфат железа), препараты органического двухвалентного железа (глюконат, фумарат и др.) не характеризуются какими-то ни было побочными эффектами. Результаты также позволяют рекомендовать дополнение курса фармакотерапии ЖДА стандартными витаминно-минеральными комплексами, так как последние содержат витамины С, В2, В6, необходимых для поддержания биологических функций железа. Следует также отметить, что анемия может быть вызвана множеством других причин (например, дефицитом фолатов), так что назначение препаратов железа должно проводиться только при подтверждении клинической симптоматики ЖДА лабораторными данными (уровни гемоглобина, ферритина, трансферрина, железа).
ЛИТЕРАТУРА
1. Torshin I. Yu., Gromova O. A. Magnesium and pyri-doxine: fundamental studies and clinical practice. Nova Science, 2009. ISBN-10: 1-60741-704-9. 250 pp.
2. Ребров В. Г., Громова О. А. Витамины, макро и микроэлементы. ГеотарМед. М., 2008, 956 с.
3. Torshin I. Yu. Bioinformatics in post-genomic era: physiol-ogy and medicine. Nova Science, 2007. ISBN10: 1-60021-752-4. 350 pp.
4. Торшин И. Ю., Громова О. А., Рудаков К. В. Иерар хия взаимодействий цинка и железа: физиологичес кие, молекулярные и клинические аспекты. Трудный пациент, #1. 2010.
5. Geisser P. Safety and efficacy of iron(III) hydroxide polymaltose complex / a review of over 25 years experi-ence. Arzneimittelforschun 2007; 57 (6A): 439—452.
6. Hart E. B., Elvehjem C. A., Kohler Go. Does liver sup-ply factors in addition to iron and copper for hemoglob-in regeneration in nutritional anemia? J. Exp. Med. 1937; 66 (2): 145—150.
7. Torshin I. Yu. Bioinformatics in post-genomic era: from molecular genetics to personalized medicine. Nova Sci ence, 2007. ISBN10: 1-60692-217-3.
8. Ignarro L. J. Nitric oxide. A novel signal transduction mechanism for transcellular communication. Hyperten sion. 1990; 16 (5): 477—483.
9. Taylor C. T., Lisco S. J., Awtrey C. S., Colgan S. P. Hypoxia inhibits cyclic nucleotide-stimulated epithelial ion transport: role for nucleotide cyclases as oxygen sen-sors. J. Pharmacol. Exp. Ther. 1998; 284 (2): 568—575.
10. Winger J. A., Marletta M. A. Expression and character-ization of the catalytic domains of soluble guanylate cy-clase: interaction with the heme domain. Biochemistry. 2005; 44 (10): 4083—4090.
11. Bezborod'ko B. N., Babak GIa. [Indices of malate and lactate dehydrogenase activity and their isoenzymes in chronic iron-deficiency anemia] Vrach Delo. 1981; (7): 41—43.
12. Branda S. S., Yang Z. Y., Chew A., Isaya G. Mitochon-drial intermediate peptidase and the yeast frataxin ho molog together maintain mitochondrial iron homeosta-sis in Saccharomyces cerevisiae. Hum Mol Genet. 1999; 8 (6): 1099—1110.
13. Reeves P. G., DeMars L. C. Copper deficiency reduces iron absorption and biological half-life in male rats. J. Nutr. 2004; 134 (8): 1953—1957.
14. Ohgami R. S., Campagna D. R., McDonald A., Flem-ing M. D. The Steap proteins are me talloreductases. Blood. 2006; 108 (4): 1388—94.
15. Chen X., Zhu C., Ji C., Zhao Y., Zhang C., Chen F., Gao C., Zhu J., Qian L., Guo X. STEAP4, a gene asso-ciated with insulin sensitivity, is regulated by several adipokines in human adipocytes. Int. J. Mol. Med. 2010; 25 (3): 361—367.
16. Zhang C. M., Chi X., Wang B., Zhang M., Ni Y. H., Chen R. H., Li X. N., Guo X. R. Downregulation of STEAP4, a highly-expressed TNF-alpha-inducible gene in adipose tissue, is associated with obesity in humans. Acta Pharmacol. Sin. 2008; 29 (5): 587—592.
17. Broker S., Meunier B., Rich P., Gattermann N., Hof-haus G. MtDNA mutations associated with sideroblastic anaemia cause a defect of mitochondrial cytochrome c oxidase. Eur. J. Biochem. 1998; 258 (1): 132—138.
18. Bannister J. V., Bannister W. H., Rotilio G. Aspects of the structure, function, and applications of superoxide dismutase. CRC Crit Rev Biochem. 1987; 22 (2): 111—80.
19. Некрасов Б. В. Основы общей химии. — М.: 1969; 857.
20. Хомченко Г. П. Пособие по химии для поступающих в вузы. М., Новая волна, 2007. С. 322.
21. Блошанский Ю. М., P. Geisser, Н. Н. Хасабов. Ане мия беременных. Consilium Medicum, Гинекология Том 08/N 2/2006.
22. He W. L., Feng Y., Li X. L., Wei Y. Y., Yang X. E. Availability and toxicity of Fe(II) and Fe(III) in Caco-2 cells. J. Zhejiang Univ Sci B. 2008 Sep.; 9 (9): 707—12.
23. Thomas C., Oates P. S. Differences in the uptake of iron from Fe(II) ascorbate and Fe(III) citrate by IEC-6 cells and the involvement of ferroportin/IREG-1/MTP 1/SLC40A1. Pflugers Arch. 2004. Jul; 448 (4): 431—7. Epub. 2004. Apr 28.
24. Астахова А. В., Лепахин В. К. Неблагоприятные по бочные реакции и контроль безопасности. Беремен ность и лекарства. Безопасность лекарств и фармако надзор, 2009. ¹ 2. С. 3—14.
25. PORRES JUAN SENABRE M.Classification of anemias and iron, copper and manganese therapy.Clin Lab (Zaragoza). 1950. Dec; 50 (297): 420—31.
26. Zhitnitskaia E. A., Griev I. G.Treatment of ancylosto-matic anemia with iron preparations in combination with vitamin B12 and trace elements. Med Parazitol (Mosk). 1966. Jul—Aug; 35 (4): 407—11.
27. Casparis D., Del Carlo P., Branconi F., Grossi A., Mer-ante D., Gafforio L. Effectiveness and tolerability of oral liquid ferrous gluconate in iron-deficiency anemia in pregnancy and in the immediate post partum period: comparison with other liquid or solid formulations con-taining bivalent or trivalent iron. Minerva Ginecol. 1996. Nov; 48 (11): 511—8. Italian.
28. Родионова Н. А., Шабаева Н. B. Опыт применения препарата Тотема при предоперационной подготовке у женщин с железодефицитной анемией. Городская кли ническая больница 1 Перинатальный центр, г. Уль яновск (Симбирск), 2003, информационное письмо.
29. Старцева Н. В., Бурдина Л. В., Швецов М. В. Лече ние препаратом «Тотема» железодефицитной анемии при беременности. Пермская государственная меди цинская академия, г. Пермь, 2001, информационное письмо.
30. Оганян Л. Ф., Баранов А. Н. Железодефицитная анемия у беременных при лечении препаратом Тоте ма. Кафедра акушерства и гинекологии Северного го сударственного медицинского университета, Архан гельская областная клиническая больница, информа ционное письмо, 2002.
Комментарии
ПРАКТИКА ПЕДИАТРА



