Экспериментальное изучение противовирусной активности Ингавирина в отношении аденовируса человека
Статьи Опубликовано в журнале:«АНТИБИОТИКИ И ХИМИОТЕРАПИЯ»; № 55; 2010; стр. 9-10.
В. В. Зарубаев1, А. В. Слита1, А. К. Сироткин1, В. Е. Небольсин2, О. И. Киселев1
2 ОАО «Валента Фарм», Москва
Изучены противовирусные свойства препарата Ингавирин® в отношении респираторного вируса человека — аденовируса 5 типа при использовании культуры клеток НЕр-2. При концентрации Ингавирина® 1000, 100 и 10 мкг/мл формируется вирусное потомство с пониженной инфекционной способностью (в 250, 100 и 10 раз соответственно). Электронно-микроскопический анализ инфицированных клеток подтвердил способность Ингавирина® нарушать нормальный морфогенез аденовируса.
Ключевые слова: Ингавирин, аденовирус человека 5 типа, противовирусная активность, электронно-микроскопический анализ, культура клеток Нер-2.
Experimental Study of Ingavirin® Antiviral Activity Against Human Adenovirus
V. V. Zarubaev, A. V. Slita, A. K. Sirotkin, V. E. Nebolsin, O. I. Kiselev
Research Institute of Influenza, North-West Branch of the Russian Academy of Medical Sciences, St.Petersburg Valenta Farm, Moscow
Antiviral properties of Ingavirin® were investigated in the Hep-2 cell culture with respect to the human respiratory tract virus (type 5 adenovirus). In concentrations of Ingavirin® of 1000, 100 and 10 mcg/ml the generated posterity showed lower infective capacity (by 250, 100 and 10 times respectivelly). The electron microscopy of the infected cells confirmed the Ingavirin® ability to disturb the adenovirus normal morphogenesis.
Key words: Ingavirin® , human type 5 adenovirus, antiviral activity, electron microscopy, Hep-2 cell culture.
Введение
Несмотря на успехи в области вакцинации, химиопрофилактики и химиотерапии респираторных вирусных инфекций человека, эти заболевания по-прежнему занимают ведущее место в структуре инфекционной патологии человека. Помимо гриппа, существенную долю респираторных инфекций составляют заболевания, вызванные вирусами других семейств: корона-, адено- и парамиксовирусами (респираторно-синцитиальный вирус и вирус парагриппа). В этиологической структуре острых респираторных вирусных заболеваний (ОРВЗ) аденовирусы человека занимают значимую позицию. Аденовирусные заболевания (АВЗ) распространены повсеместно и регистрируются в течение всего года с подъемом в зимне-весенний период, вызывая фарингиты, рино - фаринготонзиллиты, фарингоконъюнктивальную лихорадку, эпидемический керато-конъюнктивит, бронхиты и пневмонию. Отдельные случаи, нозокомиальные и локальные вспышки заболевания часто наблюдаются в закрытых детских и взрослых коллективах. АВЗ вызываются большой группой аденовирусов, из которых в патологии человека наибольшее значение имеют типы 3, 4, 5, 7, 8, 14, 21. Аденовирус 5 типа относится к латентным вирусам, который персистируя в лимфоидной ткани миндалин и аденоид в течение длительного времени, вызывает ОРВЗ у взрослых.
Активность в отношении аденовируса была показана in vitro для аналогов нуклеотидов [1, 2]. Рибавирин, используемый для терапии РС-вирусной инфекции, оказался эффективен [3, 4], частично эффективен [5] или не имел эффективности [6, 7] в отношении аденовирусной инфекции. Препарат Ганцикловир, эффективный при цитомегаловирусной инфекции, проявляет эффективность в отношении аденовируса как in vitro, так и in vivo [8, 9]. На основе (S)-1-(3-гидрокси-2-фосфонилметоксипропил) цитозина компанией Gilead Sciences был разработан препарат Цидофовир с широким спектром противовирусной активности [10—15]. Рибавирин, ганцикловир и цидофовир были с большим или меньшим успехом использованы для терапии аденовирусных патологий человека, включая гепатиты, циститы и пневмонии при иммунодефицитных состояниях у реципиентов органов [3, 4, 6, 9]. При этом выраженные побочные действия и способность аденовируса к формированию устойчивых штаммов ограничивают использование этих препаратов [13, 16, 17].
Ряд соединений других классов, таких как липиды [18], акридоны [19], имидазохинолинамины [20], как и другие аналоги нуклеозидов [1, 2, 21—24] также проявляют антиаденовирусную активность. Рост вируса в клеточной культуре может быть ингибирован катехинами зеленого чая, хотя и в высоких концентрациях [25].
Препараты интерферона, применяемые для лечения респираторных инфекций, эффективны в основном при профилактическом применении и не могут считаться основным средством терапии при тяжёлых случаях инфекции. Аденовирусы обладают эффективными механизмами подавления интерферон-индуцированного противовирусного каскада реакций, вследствие чего устойчивы к действию интерферона и его индукторов [26].
Таким образом, весьма актуальной является проблема поиска и разработки новых противовирусных препаратов, не обладающих побочными эффектами и возможно более широкого спектра действия, включая эффективность в отношении аденовируса. Ранее была показана активность препарата Ингавирин® (2-(имидазол-4-ил) этанамид пентандиовой-1,5 кислоты) в отношении вируса гриппа. Ингавирин® является низкомолекулярным аналогом эндогенного пептидоамина, выделяемого из морского моллюска Aplysia californica. Как в опытах in vitro [27], так и in vivo на модели гриппозной инфекции [28] было показано, что препарат способен ограничивать репликацию вируса в культуре клеток, а также обладает высокой протективной активностью при экспериментальном гриппе у мышей.
Цель настоящего исследования — оценка противовирусных свойств препарата Ингавирин® в отношении другого респираторного вируса человека — аденовируса 5 типа.
Материал и методы
Препараты. В работе использовали субстанцию препарата Ингавирин® в виде белого кристаллического порошка. Аликвоты препарата разводили в среде для клеточных культур Игла MEM (БиолоТ, Санкт-Петербург, кат.# 1.3.3).
Вирусы и клетки. В работе был использован аденовирус человека 5 типа из коллекции вирусных штаммов НИИ гриппа. Вирус культивировали в клетках HEp-2.
Изучение противовирусной активности препаратов. Для изучения инфекционности вирусного потомства клетки инкубировали с исследуемым препаратом в течение 1 ч, после чего в культуральную жидкость добавляли вирус (1000 CTD50 на клетку) и инкубировали в течение 1 ч. Несвязавшийся вирус отмывали 2 раза по 5 мин средой MEM и добавляли в лунки культуральную среду с препаратом в тех же концентрациях. Вирус культивировали в течение 24 ч, клетки соскабливали из лунок и титровали инфекционность вирусного потомства. Для этого из вируссодержащего материала готовили серию десятикратных разведений от 10-1 до 10-6 на среде MEM. Заражённые клетки культивировали при 37°С в атмосфере 5% СО2 в газопроточном инкубаторе Sanyo-175. Срок культивирования составлял 72 ч, после чего клетки использовали для микротетразолиевого теста (см. ниже).
Количество жизнеспособных клеток оценивали при помощи микротетразолиевого теста. Для этого клетки промывали 2 раза по 5 мин фосфатно-солевым буфером, и в лунки планшетов добавляли по 100 мкл раствора (0,5 мг/мл) 3-(4,5-диметилтиазолил-2) 2,5-дифенилтетразолия бромида (ICN Biochemicals Inc., Aurora, Ohio) на физиологическом растворе. Клетки инкубировали при 37°С в атмосфере 5% СО2 в течение 2 ч и промывали в течение 5 мин фосфатно-солевым буфером. Осадок растворяли в 100 мкл на лунку ДMСО, после чего оптическую плотность в лунках планшетов измеряли на многофункциональном ридере Victor 1420 (Perkin Elmer, Finland) при длине волны 535 нм. По результатам теста оценивали долю погибших клеток в контрольных лунках и лунках с препаратами.
Электронно-микроскопические исследования. Опытные культуры обрабатывали Ингавирином® в концентрации 1000 мкг/ мл за 1 ч до заражения вирусом(1000 CTD50 на клетку). Через 18 и 36 ч после заражения контрольные и опытные культуры клеток фиксировали 1,5% раствором глутаральдегида на среде DMEM (рН 7,2), центрифугировали 20 мин при 2000 об./мин и фиксировали 2,5% раствором OSO4. Клетки обезвоживали ацетоном в возрастающей концентрации и заливали в смесь эпон/аралдит. Ультратонкие срезы, полученные на ультрамикротоме Ultracut (Reichert, Austria), контрастировали уранилацетатом и цитратом свинца и просматривали на электронном микроскопе JEM-100S (JEOL, Japan) при инструментальном увеличении 5000—50000. Фотосъёмку производили на пленку ФТ-4ШД.
На полученных фотографиях проводили подсчёт вирионов, имеющих полноценную электронноплотную сердцевину и не имеющих её. Подсчитывали количество внутриядерных вирионов в пересчёте на одну клетку, а также процентное содержание полноценных и неполноценных вирионов в каждом случае. В каждой пробе исследовали 15—20 клеток.
Статистическая обработка данных. Статистическую обработку результатов (расчёт средних значений и стандартных отклонений, а также расчёт 50% эффективных доз при помощи линейной регрессии) проводили при помощи программы Microsoft Excel. Достоверность отличий оценивали по критерию Стьюдента. Достоверными считали различия между группами, если параметр «р» не превышал 0,05.
Результаты и обсуждение
Влияние Ингавирина® на репликацию аденовируса в культуре клеток. В проведённых экспериментах было оценено влияние Ингавирина® на инфекционность вирусной популяции, сформированной в клетках. С этой целью вирус культивировали в течение 24 ч в присутствии Ингавирина® в различных концентрациях, после чего проводили титрование инфекционности вирусного потомства во втором пассаже. Данные по влиянию Ингавирина® на инфекционность аденовируса приведены в табл. 1.
Таблица 1.
Репликация аденовируса в клетках HEp-2 в присутствии Ингавирина®
| Разведение вируса | Оптическая плотность (1000XOD535) при концентрации Ингавирина®, мкг/мл | |||
| 1000 | 100 | 10 | 0 (контроль вируса) | |
| 10-1 | 52±1 | 48±1 | 53±3 | 58±2 |
| 10-2 | 67±8 | 65±9 | 61±4 | 58±2 |
| 10-3 | 459±9 | 464±17 | 126±7 | 63±3 |
| 10-4 | 461±5 | 514±14 | 406±11 | 82±4 |
| 10-5 | 441±3 | 506±11 | 513±10 | 390±41 |
| 10-6 | 441±9 | 471±13 | 441±9 | 393±27 |
| 10-7 | 416±6 | 436±12 | 457±15 | 316±19 |
| Титр вируса, lgEID50 | 2,3 | 2,7 | 3,7 | 4,7 |
| Контроль клеток | 356±6 | |||
Как следует из представленных данных, культивирование аденовируса в присутствии Ингавирина® приводило к формированию вирусного потомства, обладающего пониженной инфекционной активностью. Степень подавления этой активности напрямую зависела от использованной концентрации препарата, и при концентрации Ингавирина® 1000, 100 и 10 мкг/мл составляла 2,4; 2,0 и 1,0 порядка (250, 100 и 10 раз) соответственно.
Ультраструктурные исследования. Как было показано в ходе ультраструктурных исследований, клетки HEp-2, инфицированные Ad5, через 18 ч после инфицирования были увеличены в размерах. Клеточные органеллы в цитоплазме выглядели интактными. В самой цитоплазме начинались процессы вакуолизации, типичные для цитодеструктивного действия аденовируса. Ядерный хроматин располагался вдоль внутреннего слоя ядерной мембраны, в кариоплазме наблюдалось формирование специфических аденовирусных включений, находящихся на ранних стадиях развития. В составе включений отчётливо различались центральные зоны умеренной электронной плотности, зоны межхроматиновых гранул, имеющие мелкоглыбчатую гранулированную структуру, и электронноплотные компактные кольца по периферии включений (рис. 1). В ядрах клеток отмечались вирионы характерной икосаэдрической формы.
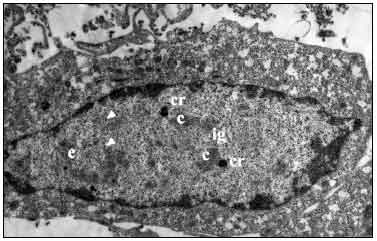
Рис. 1. Вакуолизация цитоплазмы и специфические аденовирусные включения в ядре клетки HEp-2 через 18 ч после инфицирования аденовирусом 5 типа.
C — центральные зоны; ig — интерхроматиновые гранулы; cr — компактные кольца. Вирионы аденовируса указаны стрелками. Ув. 6 000.
В клетках, инфицированных аденовирусом в присутствии Ингавирина® , признаки ранних стадий вирусной репликации были сходными с таковыми в клетках контрольной культуры, не отличаясь от них ни качественно, ни количественно. Как и в контрольных клетках, в кариоплазме наблюдалось формирование специфических аденовирусных включений, в которых отчетливо различались центральные зоны умеренной электронной плотности, зоны межхроматиновых гранул и электронноплотные компактные кольца по периферии включений (рис. 2). В ядрах клеток отмечались немногочисленные вирионы аденовируса.

Рис. 2. Аденовирусные включения в ядре клетки HEp-2 через 18 ч после инфицирования аденовирусом 5 типа в присутствии Ингавирина®.
C - центральные зоны; ig - интерхроматиновые гранулы; cr — компактные кольца. Вирионы аденовируса указаны стрелками. Ув. 6 000.
Таким образом, морфогенез собственно аденовирусной инфекции в присутствии Ингавирина® не отличался принципиально от контрольных показателей. Для более полной характеристики репродукции вируса был проведён подсчёт количества вирионов в пересчёте на одну клетку, а также подсчитана доля морфологически неполноценных вирусных частиц (табл. 2).
Таблица 2.
Морфологический состав популяции аденовируса человека 5 типа, сформировавшейся в условиях применения Ингавирина®
| Группа опыта | Количество вирусных частиц в пересчёте на клетку | |||
| с сердцевиной | без сердцевины | |||
| шт. | % | шт. | % | |
| Ингавирин® | 12±2 | 45±9* | 16±3 | 55±9* |
| Контроль без препаратов | 18±3 | 68±4 | 9±2 | 32±4 |
Как видно из представленных результатов, при подсчёте доли морфологически полноценных и дефектных вирионов были обнаружены статистически значимые различия. Так, через 18 ч после инфицирования клеток использование Ингавирина® повышало процент морфологически неполноценных вирионов с 32 до 55%.
Те же тенденции сохранялись и на более позднем сроке после инфицирования (36 ч). Количество вирусных частиц в ядрах клеток многократно возрастало, и его подсчёт, учитывая выраженное цитодеструктивное действие вируса на этой стадии, не отражал адекватно динамику вирусной репродукции. Качественно состав вирусной популяции на этой стадии представлен на рис. 3. На рисунке видно, что применение Ингавирина® существенно повышает долю незрелых вирионов, не содержащих сердцевины (рис. 3).
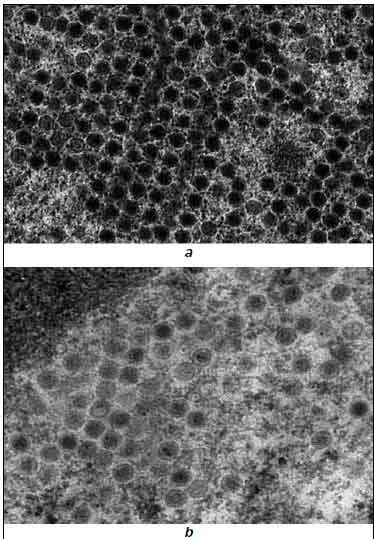
Рис. 3. Структура популяции вирионов аденовируса в клетках HEp-2 в контрольной (а) и обработанной Ингавирином® (b) культуре через 36 ч после инфицирования. Ув. 30000.
Полученные результаты можно трактовать, как способность Ингавирина® влиять, прямо или опосредованно, на процессы упаковки вирусного генома в капсиды [29—31].
Главное отличие клеток, заражённых аденовирусом в присутствии Ингавирина® от контрольной культуры без препарата, заключалось в отсутствии признаков вирусиндуцированной деградации клеток даже на дальних сроках эксперимента. Несмотря на наличие развитых внутриядерных включений и морфологически полноценных вирионов в этих клетках, цитоплазма их не содержала повышенного количества вакуолей и других цитопатогенных проявлений. На рис. 4 для наглядности представлены клетки через 36 ч после инфицирования в присутствии и без Ингавирина® . Полученные нами данные согласуются, таким образом, с ранее опубликованными результатами об активности Ингавирина® в отношении аденовируса [32].
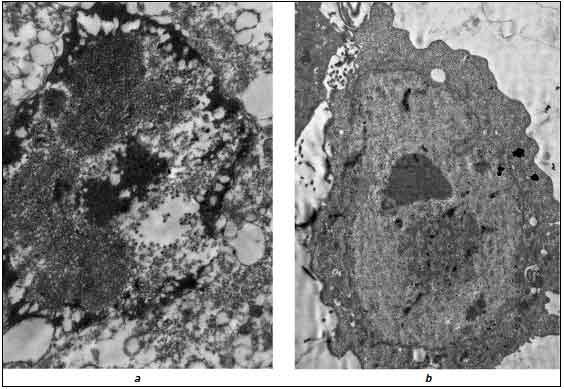
Рис. 4. Клетки HEp-2 через 36 ч после инфицирования аденовирусом 5 типа в отсутствии (а) и в присутствии (b) Ингавирина®.
а — деградация ядра и цитоплазмы, многочисленные вакуоли в цитоплазме, нарушение целостности ядерной мембраны и вирионы потомства в ядре; b — интактная цитоплазма и вирусные включения в клеточном ядре. Ув. 10 000 (a), 5000 (b).
В целом полученные данные позволяют говорить о наличии у Ингавирина® эффективного механизма защиты клеток от аденовирусной инфекции, заключающегося в способности нарушать нормальный морфогенез аденовируса, снижая тем самым инфекционность вирусного потомства.
ЛИТЕРАТУРА 1. Das A., Trousdale М. D., Ren S, Lien Е. J. Inhibition of Herpes simplex virus type 1 and adenovirus type 5 by heterocyclic Schiffbases of amino-hydroxyguanidine tosylate. Antiviral Res 1999; 44: 3: 201—208.
2. De Clercq Е, Sakuma Т., Baba M. et al. Antiviral activity of phospho-nylmethoxyalkyl derivatives of purines and pyrimidines. Аntiviral Res 1987; 8: 5—6: 261—272.
3. Arav-Boger R, Echavarria M., Forman M. et al. Clearance of adenoviral hepatitis with ribavirin therapy in a pediatric liver transplant recipient. Pediatr Infect Dis J 2000; 19: 11: 1097—1100.
4. Shetty A. K, Gans H. A., So S. et al. Intravenous ribavirin therapy for adenovirus pneumonia. Pediatr Pulmonol 2000; 29: 1: 69—73.
5. Gavin P. J., Katz B. Z. Intravenous Ribavirin treatment for severe adenovirus disease in immunocompromised children. Pediatrics 2002; 110: 1 pt 1: 1—8.
6. Bordigoni P., Carret A. S, Venard V. et al. Treatment of adenovirus infections in patients undergoing allogeneic hematopoietic stem cell transplantation. Clin Infect Dis 2001; 32: 9: 1290—1297.
7. La Rosa A. M, Champlin R. Е, Mirza M. et al. Adenovirus infections in adult recipients of blood and marrow transplants. Clin Infect Dis 2001; 32: 6: 871—876.
8. Trousdale M. D, Goldschmidt P. L, Nobrega R. Evaluation of ganciclovir against human adenovirus type 5 infection in cell culture and cotton rat eyes. Antiviral Res 1993; 20: Suppl. 1: 48.
9. Chen F. Е, Liang R. H, Lo J. Y. et al. Treatment of adenovirus-associated haemorrhagic cystitis with ganciclovir. Bone Marrow Transplant 1997; 20: 11: 997—999.
10. Hillenkamp J., Reinhard Т., Ross R. S. et al. The effect of cidofovir 1% with and without cyclosporin A 1% as а topical treatment of acute adenoviral keratoconjunctivitis: а controlled clinical pilot study. Ophtalmology 2002; 109: 5: 845-850.
11. Hoffman J. A., Shah A. J., Ross L. A., Kapoor N. Adenoviral infections and а prospective trial of cidofovir in pediatric hematopoietic stem cell transplantation. Biol Blood Marrow Transplant 2001; 7: 7: 388—394.
12. Romanowski E. G, Yates K. A., Gordon Y. J. Antiviral prophylaxis with twice daily topical cidofovir protects against challenge in the adenovirus type 5/New Zealand rabbit ocular model. Antiviral Res 2001; 52: 3: 275—280.
13. Romanowski E. G, Gordon Y. J., Araullo-Cruz T. et al. The antiviral resistance and replication of cidofovir-resistant adenovirus variants in the New Zealand white rabbit ocular model. Invest Ophtalmol Vis Sci 2001; 42: 8: 1812—1815.
14. Safrin S, Cherrington J., Joffe H. S. Cidofovir. Review of current and potential clinical uses. Adv Exp Med Biol 1999; 458: 111—120.
15. Hitchcock M. J. M., Jaffe H. S, Martin J. C., Stagg R. J. Cidofovir, a new agent with potent anti-herpesvirus activity. Antivir Chem Chemother 1996;7: 3: 115—127.
16. Gordon Y. J., Naesens L., De Clercq E. et al. The treatment of adenoviral conjunctivitis with topical cidofovir. Cornea. 1996;15: 5: 546.
17. Coca S., Perazella M. A. Rapid communication: acute renal failure associated with tenofovir: evidence of drug-induced nephrotoxicity. Am J Med Sci 2002; 324: 6: 342—344.
18. Kaneko H, Kato K, Mori S, Shigeta S. Antiviral activity of NMSO3 against adenovirus in vitro. Antiviral Res 2001; 52: 3: 281—288.
19. Zarubaev V. V., Slita A. V., Krivitskaya V. Z. et al. Direct antiviral effect of cycloferon (10-carboxymethyl-9-acridanone) against adenovirus type 6 in vitro. Antiviral Res 2003; 58: 2: 131—137.
20. Rautenschlein S., Miller R. L., Sharma /. M. The inhibitory effect of the imidazoquinolinamine S-28828 on the pathogenesis of a type II adenovirus in turkeys. Antiviral Res 2000; 46: 3: 195—205.
21. Naesens L., Lenaerts L., Andrei G. et al. Antiadenovirus activities of several classes of nucleoside and nucleotide analogues. Antimicrob Agents Chemother 2005; 49: 1010—1016.
22. Mentel R, Kinder M., Wegner U. et al. Inhibitory activity of 3-fluoro-2-deoxythymidine and related nucleoside analogues against adenoviruses in vitro. Antiviral Res 1997; 34: 3: 113—119.
23. Zarubaev V. V., Slita A. V., Sukhinin V. P. et al. Effect of 6-azacytidine on the course of experimental adenoviral infection in newborn Syrian hamsters. J Chemother 2007; 19: 1: 44-51.
24. Alexeeva I., Dyachenko N, Nosach L.et al. 6-Azacytidine — compound with wide spectrum of antiviral activity. Nucleosides Nucleotides Nucleic Acids. 2001; 20: 4—7: 1147—1152.
25. Weber J. M., Ruzindana-Umunyana A., Imbeault L., Sircar S. Inhibition of adenovirus infection and adenain by green tea catechins. Antiviral Res 2003; 58: 2: 167—173.
26. Zhang Y, Schneider R. J. Adenovirus inhibition of cellular protein synthesis and the specific translation of late viral mRNAs. Semin Virol 1993; 4: 229-236.
27. Логинова С. Я., Борисевич С. В., Лыков М. В. и др. Изучение эффективности Ингавирина® in vitro в отношении «мексиканского» пандемического подтипа H1N1 вируса гриппа А, штаммы А/California/04/2009 и A/California/07/2009. Антибиотики и химиотер 2009; 3—4: 15—17.
28. Логинова С. Я., Борисевич С. В., Максимов В. А. и др. Изучение терапевтической эффективности нового отечественного препарата Ингавирин в отношении возбудителя гриппа А (H3N2). Там же 2008; 53: 11/12: 27—30.
29. Xing L, Tikoo S. K. Packaging of viral RNAs in virions of adenoviruses. Virol J 2009; 5: 6: 16.
30. Erturk E, Ostapchuk P., Wells S. I. et al. Binding of CCAAT displacement protein CDP to adenovirus packaging sequences. J Virol 2003; 77: 11: 6255—6264.
31. Ostapchuk P., Hearing P. Regulation ofadenovirus packaging. Curr Top Microbiol Immunol. 2003; 272: 165—185.
32. Логинова С. Я, Борисевич С. В., Максимов В. А. и др. Изучение эффективности Ингавирина® in vitro в отношении возбудителя аденовирусной инфекции. Антибиотики и химиотерапия, 2009, 54; 7—8.
Комментарии
ПРАКТИКА ПЕДИАТРА



