Роль меди и марганца в метаболизме железа
Статьи Опубликовано в журнале:«ВРАЧ»; Фармакология; 12; 2013; стр. 47-52. Е. Семенова1, М. Кунина2,
Н. Стуклов1, доктор медицинских наук, профессор
2ООО Клиника «Будь здоров», Москва
В метаболизме железа в организме человека принимают участие 22 марганцевых белка и 9 купроэнзимов. Монотерапия железом не учитывает взаимодействия между микроэлементами и поэтому может сопровождаться повышенным числом побочных эффектов, а в случае дефицита меди вызвать перегрузку организма железом либо рефрактерность к лечению. Применение комбинации железо + медь + марганец позволяет избежать подобных результатов.
Ключевые слова: лечение анемии, железа глюконат, железа сульфат, дефицит железа, дефицит меди, глюконат меди, глюконат марганца, Тотема, Сорбифер дурулес.
Role Of Copper And Manganese In Iron Metabolism
E. Semenova1, M. Kunina2,
Professor N. Stuklov1, MD
2OOO Be Healthy Clinic, Moscow
Twenty-two manganese proteins and 9 cuproenzymes are involved in human iron metabolism. Iron monotherapy fails to take into account trace element interactions and may be therefore accompanied by the higher rates of side effects, and, in case of copper deficiency, may cause the body's iron overload or treatment refractoriness. The use of a combination of iron, copper, and manganese allows such results to be avoided.
Key words: anemia treatment, ferrous gluconate, ferrous sulfate, iron deficiency, copper deficiency, copper gluconate, manganese gluconate, Totema, Sorbifer durules.
Железо выполняет в организме ряд очень важных функций — захват, транспорт кислорода (гемоглобин), хранение кислорода (миоглобин), выработка энергии в клетках (цитохром с-оксидаза). Железо участвует в механизмах иммунитета (ионы Fe2+, лактоферрин), антиоксидантной защите (пероксидаза, каталаза), синтезе ДНК (коэнзимредуктаза рибонуклеотидов), в формировании соединительной ткани (лизилгидроксилаза и пролилгидроксилаза-1 отвечают за стабилизацию коллагена и синтез хондроитинсульфат протеогликанов). На сегодня насчитывается около 100 железосодержащих белков [1—4].
Количество железа в организме в норме чрезвычайно стабильно и определяется равновесием между его поступлением и потерями. Всего в организме взрослого человека содержится от 3 до 5 г железа, причем до 70% его инкорпорировано в гемоглобин, приблизительно 10% находится в миоглобине и железосодержащих энзимах, остальное сохраняется в медленно обменивающемся пуле в составе ферритина — ФТ (его называют также пулом накопления) [3, 4]. Плазматический транспорт включает трансферриновое железо и составляет приблизительно 0,1%. Ежедневные потери железа чрезвычайно малы, порядка 1 мг в день, и являются результатом слущивания эпителия, потерь с желчью; меньше железа выводится с мочой. У женщин репродуктивного возраста к этому добавляются ежемесячные потери железа в связи с менструальными кровотечениями. Компенсация этих потерь имеет важнейшее значение и тщательно регулируется, поскольку человеческий организм не имеет возможности увеличить экскрецию железа в случае его избыточного поступления [3—5].
Роль меди и марганца в абсорбции железа в кишечнике и утилизации в тканях
В гомеостазе железа принимают участие 9 медьсодержащих ферментов и 22 марганецзависимых белка. Важная роль в метаболизме железа отводится таким купроэнзимам, как дуоденальный цитохром b (Dcyt b), гефестин, эндосомальные ферриредуктазы STEAP и церулоплазмин, которые осуществляют взаимопревращения Fe2+ и Fe3+, необходимые при входе железа в клетки, выходе железа в кровоток и дальнейшем связывании с трансферрином [2, 4]. Всасывание железа происходит преимущественно в верхних отделах тонкой кишки. Наибольшей биодоступностью (до 30%) обладает железо, входящее в состав гема (мясные продукты); оно усваивается в неизменном виде с помощью пиноцитоза, предварительно связавшись с мембранным белком-переносчиком гема (НСР1). В образовавшейся эндосоме под действием гемоксигеназы (Hox) железо высвобождается в цитоплазму в так называемый транзитный пул либо связывается с ФТ [3, 4]. Этот путь является основным для поступления железа из пищи. Негемовое железо представлено нерастворимыми гидроокисями и потому требует активного транспорта через неспецифические марганецзависимые транспортеры двухвалентных металлов (ДМТ1) с участием медьзависимого дуоденального цитохрома b, который восстанавливает окисное железо Fe3+ до растворимой закисной формы Fe2+. Биодоступность негемового железа составляет всего 1—3%. Далее через базальную мембрану энтероцита также в двухвалентном состоянии железо попадает в кровоток через мембранный белок ферропортин. Ферропортин связан с купроэнзимом гефестином, который является мембранной феррооксидазой и окисляет железо для связывания с переносчиком трансферрином, как показано на рис. 1 [2—5].
Рис. 1. Всасывание железа в кишечнике: медь в составе дуоденального цитохрома (Dcyt b), гефестина и церулоплазмина участвует в механизме активного транспорта железа и связывании железа с трансферрином; через марганецзависимые транспортеры двухвалентных металлов (ДМТ1) железо попадает внутрь энтероцита 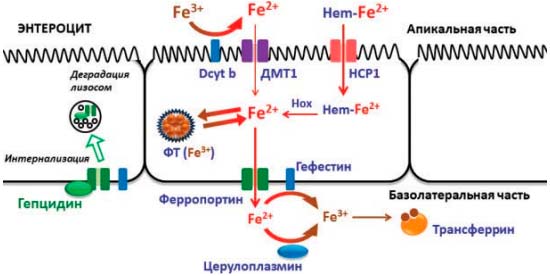
В сутки на нужды эритропоэза требуется порядка 20—25 мг железа. Таким образом, железо поступающее из энтероцитов, составляет всего около 5%, остальные 95% поступают из системы мононуклеарных макрофагов, осуществляющих реутилизацию старых эритроцитов [4, 5].
Продолжительность жизни эритроцита в среднем — 125 сут. Стареющие клетки фагоцитируются макрофагами, и в образовавшейся фаголизосоме под действием гемоксигеназы высвобождаются ионы Fe2+, которые через марганецзависимые белки ДМТ1 выходят в цитоплазму, далее через ферропортин — в кровоток или накапливаются в ФТ. Железо из макрофагов, вышедшее в кровоток, окисляется церулоплазмином для связывания с трансферрином (рис. 2).
Рис. 2. Реутилизация железа макрофагами ретикулоэндотелиальной системы: марганец в составе белка ДМТ1 способствует выходу железа из фаголизосомы в цитоплазму клетки 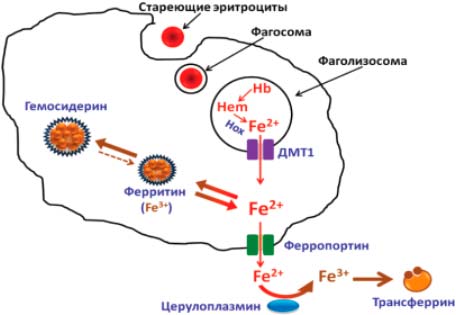
Фракция железа, не предназначенная для костного мозга, делится между различными местами накопления, представленными макрофагами, и в основном гепатоцитами, особенно чувствительными к перегрузке железом, а также доставляется в клетки разных органов. Комплекс железо— трансферрин захватывается рецептором-1 трансферрина (TfR1), затем пиноцитируется (рис. 3). В результате он оказывается внутри лизосом, куда с помощью АТФ-насоса нагнетаются ионы Н+, и происходит закисление среды. В кислой среде комплекс трансферрин—Fe3+ диссоциирует, далее Fe3+ восстанавливается эндосомальной Cu-НАДН-зависимой ферроокидазой (Fe-R) до растворимого Fe2+ и через белки ДМТ1 попадает в цитоплазму в лабильный (транзитный) пул (ЛПЖ), откуда транспортируется в места синтеза железосодержащих белков либо попадает в пул накопления. В транзитном пуле железо находится в виде Fe2+, поскольку несвязанные ионы Fe3+ неустойчивы при физиологичных рН и быстро образуют нерастворимые в воде оксиды и гидроокиси [2—5].
Рис. 3. Рецепторопосредованный (TfR) захват железа клеткой из трансферрина: медь в составе эндосомальной НАДН-зависимой редуктазы (Fe-R) восстанавливает железо для выхода из эндосомы в цитоплазму клетки через марганецзависимый транспортер (ДМТ1) 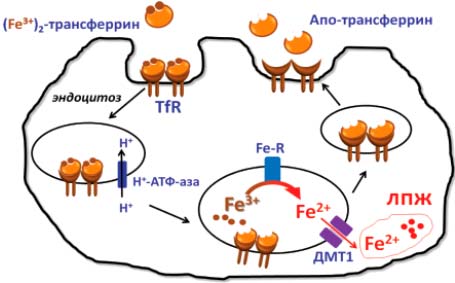
Помимо упомянутого марганецзависимого белка ДМТ1, играющего ключевую роль в транспорте железа внутрь клеток, всего на биологические функции железа влияют 22 марганцевых энзима (серинтреонинфосфатазы, гуанилатциклазы, ион-транспортеры, малатдегидрогеназа, митохондриальная пептидаза). Из них наиболее изучена митохондриальная пептидаза, при участии которой созревают белковые молекулы, в том числе транспортеры железа, и регулируется уровень железа в митохондриях, где происходит последняя стадия синтеза гема [2].
Роль меди в регуляции гомеостаза железа
Регуляция абсорбции железа осуществляется на уровне базальной мембраны энтероцита с помощью гепцидина. Этот небольшой противомикробный белок (25 аминокислот) вырабатывается в основном в печени. Его синтез активируется при высоких уровнях запасов железа, высокой концентрации железа в сыворотке, а также при инфекции и воспалении под действием воспалительных цитокинов, таких как интерлейкин-6. В результате связывания гепцидина с ферропортином индуцируется его интернализация в клетку с последующей деградацией в лизосомах (см. рис. 1). Таким образом, железо оказывается запертым внутри клетки и утрачивается в процессе естественной десквамации кишечного эпителия. Аналогичным образом гепцидин регулирует выход железа в кровоток из макрофагов и других клеток. В случае высокой активности эритропоэза (независимо от состояния запасов железа), при дефиците железа или гипоксии выработка гепцидина подавляется, и железо через ферропортин попадает в кровоток, где после окисления гефестином или церулоплазмином связывается с трансферрином и переносится в костный мозг [4—6].
Интересны данные F. Martin и соавт. о роли меди в активации эритропоэза путем стабилизации выработки индуцированного гипоксией фактора (HIF1), который контролирует экспрессию гена эритропоэтина [7]. Регуляция выработки HIF1 осуществляется семейством пролилгидроксилазных доменов (PHD) ферментов, которые выполняют функции датчиков кислорода и Fe2+ в клетках. В условиях нормоксии PHD гидроксилируют а-субъединицу HIF, что приводит к ее деградации. При снижении парциального давления кислорода в клетках (гипоксии) активность PHD ферментов снижается, что приводит к накоплению ядерного HIF и индукции генов-мишеней. У млекопитающих известны 3 HIF-пролилгидроксилазы, кодируемые независимыми генами, PHD1, PHD2, PHD3. При этом если PHD2 и PHD3 прекращают индуцированную гипоксией HIF-транскрипцию при реоксигенации, то PHD1 является кислороднезависимой [8]. В экспериментах F. Martin и соавт. было показано, что медь при нормальном парциальном давлении кислорода стабилизирует ядерный HIF1 и ингибирует пролил-4-гидроксилирование. Основываясь на спектрометрических и структурных исследованиях 2-оксоглуторат-зависимых диокигеназ, показывающих, что Cu2+ может заменить Fe2+ в активном центре в этом классе энзимов, можно предположить, что ионы Cu2+ связываются с активным центром PHD со значительно более высоким сродством, чем ионы Fe2+, однако в эксперименте F. Martin ионы меди тормозили гидроксилирование HIF даже при избытке ионов железа. Таким образом, механизм, ответственный за регуляцию выработки HIF1 при участии ионов меди, еще неизвестен. Основными мишенями для HIF являются гены эритропоэтина (синтезируется преимущественно в почках), а также трансферрина, трансферриновых рецепторов и церулоплазмина (синтезируются в печени) [7, 8]. Увеличение уровня эритропоэтина приводит к активации эритропоэза, что ведет к снижению содержания гепцидина, увеличению всасывания железа в кишечнике и мобилизации железа из макрофагов ретикулоэндотелиальной системы. Увеличение синтеза церулоплазмина и трансферрина способствует быстрому усвоению железа в тканях, нормализации синтеза гемоглобина и других железосодержащих белков.
Различия в механизме всасывания железа из антианемических препаратов
Если для пищевого железа основным путем поступления этого микроэлемента является путь гема, то для всасывания железа из препаратов основными каналами будут белки ДМТ1. При этом препараты трехвалентного железа будут адсорбироваться путем активного транспорта, так же как негемовое железо пищи, а двухвалентные препараты железа — свободно диффундировать через каналы транспортеров двухвалентных металлов. Именно поэтому биодоступность таких препаратов, как Ферлатум, Мальтофер, Феррум Лек, Биофер, в состав которых входит трехвалентное железо, не превышает 10%, а биодоступность препаратов на основе двухвалентного железа (Тотема, Ферретаб, Сорбифер, Тардиферон, Ферро-фольгамма, Фенюльс, Актиферрин, Гемофер, Ферроградумет, Ферроплекс) составляет от 10 до 30% [13, 17, 18].
Влияние дефицита меди на адсорбцию из двухвалентных и трехвалентных препаратов
Отдельно необходимо затронуть проблемы лечения железодефицитной анемии (ЖДА) в случае ее сочетания с дефицитом меди. Поскольку до 80% всей меди находится в плазме в составе церулоплазмина, велика вероятность того, что ЖДА, развившаяся в результате хронической или острой кровопотери, будет сопровождаться дефицитом меди. Дефицит меди может возникать и у недоношенных детей, и в период интенсивного роста. По данным разных авторов, частота дефицита меди у детей дошкольного возраста — 58%, у детей школьного возраста — от 39 до 89% [10, 11]. Исследование микроэлементного состава крови юношей 17—19 лет, проживающих во Владивостоке, выявило дефицит меди в 63% случаев [12].
При лечении ЖДА в сочетании с дефицитом меди препараты трехвалентного железа будут неэффективны, поскольку в отсутствие этого микроэлемента нарушается активный транспорт железа (рис. 4). В литературе имеются данные о неэффективности применения полимальтозного комплекса гидроксида железа III (ПКГЖ), даже при длительном лечении (от 4 до14 мес), у значительного числа пациентов — 75 (31%) из 241. Последующий перевод этих пациентов на препараты двухвалентного железа приводил к быстрой нормализации гемоглобина [14, 15].
Рис. 4. При дефиците меди нарушается механизм активного транспорта железа, поэтому применение препаратов железа III может привести к рефрактерности терапии 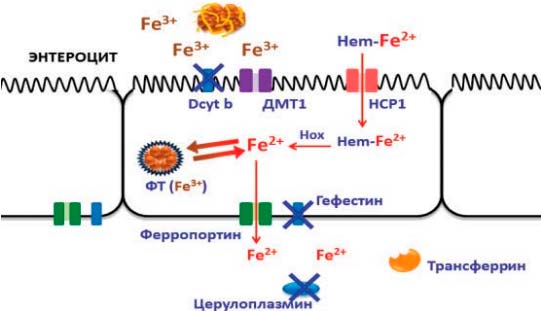
При лечении препаратами на основе двухвалентных солей железа ионы Fe2+ свободно проходят через каналы белков ДМТ1, но из-за снижения активности гефестина и церулоплазмина будет нарушена утилизация железа и возможно развитие состояния перегрузки железом с активацией свободнорадикального окисления, причем чем больше дефицит меди, тем вероятнее возникновение такого состояния (рис. 5).
Рис. 5. Применение препаратов железа II при сочетанном дефиците меди может вызвать перегрузку организма железом из-за нарушенной утилизации железа 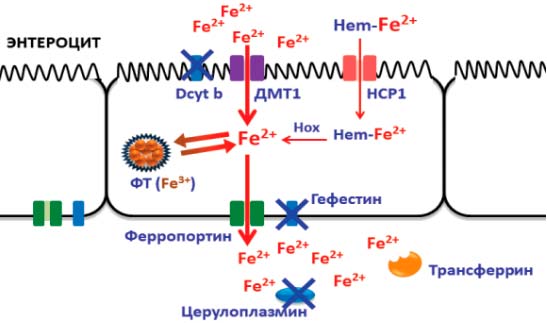
Применение комбинации железо + медь позволяет избежать подобных проблем. Однако следует помнить, что заболевания печени, сопровождающиеся снижением ее синтетической функции, могут приводить к недостатку выработки белков, связывающих медь (церулоплазмин, альбумин); в случаях наследственных заболеваний, таких как болезнь Вильсона—Коновалова, встречается обратная ситуация: избыток свободных ионов меди, и тогда применение медьсодержащих препаратов противопоказано.
Роль меди и марганца в антиоксидантнои защите
Известно, что железо способно активировать процессы образования свободных радикалов, поэтому поддержание баланса антиоксидантных молекул в клетках и плазме — необходимое условие нормального функционирования организма. Наиболее важные антиоксиданты в клетках — супероксиддисмутаза (СОД) и глутатион, в мембранах — токоферол и p-каротин, в плазме — церулоплазмин. В активном центре цитозольной СОД находится медь, а в СОД митохондрий — марганец. Содержащий до 8 ионов меди церулоплазмин инактивирует железо, связывая его с трансферрином, и, кроме того, обладает слабой СОД-активностью. Важно, что применение препаратов железа может снижать синтез Mn-СОД, так как железо конкурирует с марганцем за участок связывания на уровне транскрипции этого фермента, тогда как комбинация железо + марганец лучше удовлетворяет потребности в этом антиоксидантном ферменте [3, 9, 17].
Принимая во внимание роль меди и марганца в механизмах всасывания и утилизации железа, а также механизмы антиоксидантной защиты, мы сравнили применение при ЖДА препарата Тотема (производитель — Лаборатория Иннотек Интернасиональ), содержащего глюконат железа II, медь и марганец, и препарат Сорбифер дурулес, в состав которого входит сульфат железа II и витамин С. По данным литературы, жидкий глюконат железа в более низких дозах так же эффективен, как таблетированные формы сульфата и глюконата железа [16]. Поэтому мы применяли низкие дозы жидкого комбинированного препарата Тотема и высокие дозы таблетированного препарата Сорбифер дурулес.
Результаты сравнения эффективности жидкого глюконата железа II в комбинации с глюконатом меди и марганца и таблетированной формы с замедленным высвобождением сульфата железа II в комбинации с витамином C
В исследовании приняли участие 19 женщин с ЖДА, развившейся в результате хронической кровопотери на фоне аденомиоза и гиперплазии эндометрия. Лечение проводилось на фоне основного заболевания при продолжающейся кровопотере. Пациентки были разделены на 2 группы, сопоставимые по возрасту, тяжести заболевания и анемии. 1-я группа (11 женщин, средний возраст — 33±7 года) получала комбинацию глюконата железа + медь + марганец по 2 ампулы в день, что соответствует дозе 100 мг железа в сутки. Пациенток 2-й группы (8 женщин, средний возраст — 34±8 года) лечили комбинацией сульфата железа с витамином С по 2 таблетки в день, что соответствует дозе 200 мг железа в сутки. Результаты исследования представлены на рис. 6.
Рис. 6. Сравнение эффективности применения низких доз жидкого глюконата FeII, меди и марганца и высоких доз таблетированного сульфата FeII с витамином С при лечении ЖДА легкой степени 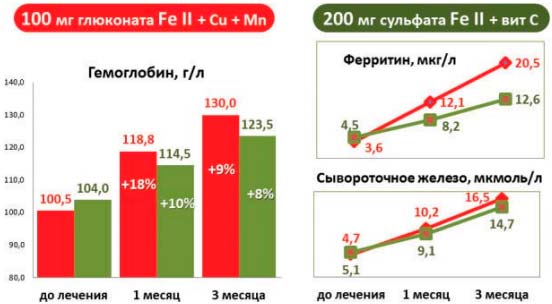
В обеих группах показатели гемоглобина и сывороточного железа через 1 и 3 мес терапии были сопоставимыми, несмотря на различие применяемых доз; при этом показатели ФТ через 3 мес лечения достоверно различались (р=0,05), причем в 1-й группе, применявшей низкие дозы жидкого глюконата железа в комбинации с медью и марганцем, они были выше (20,5±11,9 мкг/л), чем в группе, применявшей высокие дозы таблетированного сульфата железа в комбинации с витамином С (12,6±3,9 мкг/л).
Результаты сравнения переносимости жидкой формы глюконата железа ii, меди и марганца (тотема) и препаратов двухвалентного и трехвалентного железа [17]
Метаанализ 30 исследований (n=1077) показал хорошую переносимость препарата Тотема. Средняя частота нежелательных явлений составила 10,4% (n=112). Побочные явления в основном были представлены легкими, непродолжительными диспепсическими реакциями (8,5% случаев), не требующими отмены препарата. Реже встречались аллергические реакции в виде необильной сыпи (0,7% случаев) и потемнение эмали зубов (1,0%). Только 1,1% пациентов были вынуждены отказаться от дальнейшего приема препарата Тотема.
Сопоставление данных этих 30 исследований применения препарата Тотема с величинами, полученными в метаанализе 5 сравнительных исследований применения ПКГЖ (n=289) и сульфата железа II (n=208) и в метаанализе 3 сравнительных исследований применения протеин-сукцинилата железа III (n=890) и сульфата железа II (n=634), показало, что препарат Тотема обладает лучшей переносимостью (побочные эффеты в 10,4% случаев), чем сульфат железа II (в 5 исследованиях побочные эффекты возникли у 34,1% больных, в 3 — у 20,4%; р<0,005), и его переносимость достоверно не отличается от переносимости препаратов трехвалентного железа — ПКГЖ (побочные эффекты - у 14,9% больных; 0,1<р<0,25) и протеина сукцинилата железа III (у 9,4%; 0,75<р<0,9), рис. 7.
Рис. 7. Сравнение результатов исследований переносимости различных железосодержащих препаратов [17] 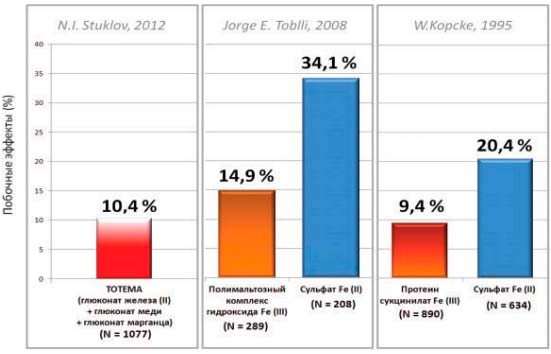
Отдельно сравнивали частоту запоров, поскольку данный побочный эффект может значительно снизить приверженность лечению, спровоцировать обострение геморроя и крайне нежелателен при беременности. Частота запоров при применении препарата Тотема (24 исследования, n=841) составила всего 0,83% (р<0,005), что в 10 раз меньше, чем при применении ПКГЖ (8,7%) и в 19 раз меньше, чем при применении сульфата железа (15,9%) [17].
Изложенное позволяет заключить, что:
Рекомендации производителя по использованию жидкой формы глюконата железа II, меди и марганца (Тотема)
Для минимизации вероятности появления нежелательных явлений при приеме препарата Тотема необходимо:
ЛИТЕРАТУРА
1. Керимкулова Н.В., Торшин И.Ю., Громова О.А. и др. Систематический анализ молекулярно-физиологических эффектов синергичного воздействия железа, марганца и меди на соединительную ткань // Гинекология. - 2012; 14 (6).
2. Громова О.А., Торшин И.Ю., Хаджидис А.К. Анализ молекулярных механизмов воздействия железа (II), меди, марганца в патогенезе железодефицитной анемии // Клин. фармакология и фармаэкономика. - 2010; 1: 1-9.
3. Владимиров Ю.А. и др. Свободные радикалы в живых системах. ВИНИТИ Итоги науки и техники / М.: Биофизика, 1991; Том 29: 1-250.
4. Cadet E., Gadenne M., Capront D. et al. Donnes recentes sur metabolisme du fer: un etat de transition // Rev. Med. In. - 2005; 26: 315-24.
5. Павлов А.Д., Морщакова Е.Ф., Румянцев А.Г. Эритропоэз, эритропоэтин, железо. Молекулярные и клинические аспекты / М.: ГЭОТАР-Медиа, 2011; 300 с.
6. Левина А.А., Казюкова Т.В., Цветаева Н.В. и др. Гепсидин как регулятор гомеостаза железа // Педиатрия. - 2008; 87 (1): 67-74.
7. Martin F., Liden T.,Katschinski D. et al. Copper-dependent activation of hypoxia-inducible factor (HIF)-1: implications for ceruloplasmin regulation // Blood. - 2005; 105 (12): 4613-9.
8. Fong G., Takeda K. Role and regulation of prolyl hydroxylase domain proteins // Cell. Death Differ. - 2008; 15 (4): 635-41.
9. Davis C., Greger J. Longitudinal changes of manganese-dependant superoxide dismutase and other indexes of manganese and iron in women // Am. J. Clin. Nutr. - 1992; 55: 747-52.
10. Иванова Н.А. Возможности коррекции микроэлементозов у детей школьного возраста // Практика педиатра. - 2011: 16-20.
11. Вильмс Е.А., Турчанинов Д.В., Турчанинова М.С. Микроэлементозы у детского населения мегаполиса: эпидемиологическая характеристика и возможности профилактики // Педиатрия. - 2011; 90 (1): 96-101.
12. Антонюк М.В., Симонова И.Н., Андрюков Б.Г. и др. Микроэлементный статус юношей Владивостока // Здоровье. Медицинская экология. Наука. -2009; 1 (36): 12-6.
13. Nielsen P., Gabbe E., Ficher R. et al. Bioavailability of iron from oral ferric polymaltose in humans // Arzneimittel-Forschung/Drug Research. - 1994; 44 (6): 743-8.
14. Ruiz-Arguelles G., Diaz-Hernandez A., Manzano C. et al. Ineffectiveness of oral iron hydroxide polymaltose in iron-deficiency anemia // Hematology. - 2007; 12 (3): 255-6.
15. Mehta B. Iron (III) hydroxide polymaltose is ineffective in treatment of iron deficiency anemia // Med. Image. - 2001; 25: 36-7.
16. Casparis D., Del Carlo P., Braconi F. et al. Effectiveness and tolerability of oral liquid ferrous gluconate in iron-deficiency anemia in pregnancy and in the immediate poste-partum period: comparison with other liquid or solid formulations containing bivalent or trivalent iron // Min. Ginecol. - 1996; 48 (11): 511-8.
17. Стуклов Н.И. Мета-анализ данных переносимости питьевой формы глюконата железа (II), меди и марганца (препарат Тотема) при лечении железодефицитной анемии у детей и взрослых // Земский врач. - 2012; 4 (15): 11-20.
18. Стуклов Н.И., Семенова Е.Н. Лечение железодефицитной анемии. Что важнее эффективность или переносимость? Существует ли оптимальное решение? // Журнал международн. мед. Гинекология. - 2013; 1 (2): 47-55.
Комментарии
ПРАКТИКА ПЕДИАТРА



