Исследование действия мексидола при "избегаемом" и "неизбегаемом" эмоциональном стрессе у мышей инбредных линий BALB/C И C57BL/6
СтатьиОпубликовано в журнале:
« Экспериментальная и клиническая фармакология » Том 67 № 6 2004
О. Ю. Кравцова, Т. А. Воронина, А. К. Сариев1
1 Лаборатории психофармакологии (руководитель - проф. Т. А. Воронина), фармакокинетики (руководитель - проф. В. П. Жердев) ГУ НИИ фармакологии им. В. В. Закусова РАМН, Москва, 125315, ул. Балтийская, 8
Изучено действие мексидола (100мг/кг внутрь) у мышей инбредных линий BALB/C и C57BL/6 на моделях эмоционального стресса ("открытое поле", "приподнятый крестообразный лабиринт", "условный рефлекс пассивного избегания"). Установлено, что препарат после однократного введения оказывает селективное анксиолитическое действие на мышей линии BALB/C - животных с "пассивным" фенотипом эмоционально-стрессовой реакции (ЭСР) и не проявляет седативного эффекта у мышей линии C57BL/6 с "активным" фенотипом ЭСР.
Ключевые слова: мексидол, инбредные животные, эмоциональный стресс
Investigation of the effects of mexidol in escapable and inescapable emotional stress in inbred BALB/C and C57BL/6 MICE
O. Yu. Kravtsova, T. A. Voronina, and A. K. Sariev
Zakusov Institute of Pharmacology, Russian Academy of Medical Sciences, Baltiiskaya ul. 8, Moscow, 125315 Russia
The effect of mexidol administered in a dose of 100 mg/kg (i.p.) was studied in BALB/C and C57BL/6 mice on various emotional stress models including open field test, elevated plus-maze test, and passive avoidance conditioned reflex Upon a single administration, the drug produces a selective anxiolytic action on BALB/C mice (experimental animals with a "passive" phenotype of the emotional stress reaction), while not inducing any sedative (tranquilizer) effect in C57BL/6 mice (belonging to the phenotype with "active" emotional stress reaction).
Введение
И генетически гетерогенной популяции известна широкая вариабельность показателя тревожности, определяемая, прежде всего, различиями в наследственно контролируемых типах реакции на стресс. В связи с этим при проведении фармакологической коррекции нарушений поведения при эмоциональном стрессе следует учитывать индивидуальные особенности [1, 9]. В лечебной практике в качестве анксиолитического средства применяется препарат мексидол (2-этил-6-метил-3-оксипиридина сукцинат) [4, 5, 7]. В эксперименте на мышах инбредных линий показано, что мексидол в тесте "открытое поле" устраняет "реакцию замирания" (freezing) у мышей BALB/C, не оказывая при этом седативного действия на мышей C57BL/6 [9, 12].
Целью настоящего исследования явилось сравнительное изучение антистрессорного действия мексидола у беспородных мышей и мышей инбредных линий BALB/C и C57BL/6 с различными врожденными типами эмоционально-стрессовой реакции (ЭСР) в условиях "избегаемого" (тесты "приподнятый крестообразный лабиринт" и "условный рефлекс пассивного избегания") и "неизбегаемого" (тест "открытое поле") эмоционального стресса.
Методы исследования
Опыты выполнены на мышах-самцах инбредных линий BALB/C и C57BL/6 и белых беспородных мышах-самцах массой 26 - 30 г, полученных из питомника Столбовая РАМН. Животных содержали в стандартных условиях вивария. Эксперименты проводили в интервале 10 - 16 ч. Водный раствор мексидола готовили заранее и вводили мышам в желудок с помощью металлического зонда за 30 мин до тестирования в дозе 100мг/кг. Контрольные животные получали равный объем дистиллированной воды.
Тест "открытое поле" (ОП) проводили по стандартной методике [2, 3]. Поведение животных оценивали в течение трех минут, регистрируя горизонтальную двигательную активность (количество пересеченных квадратов), вертикальную активность (число вставаний на задние лапы), исследовательскую активность (число обследованных отверстий), а также количество болюсов (эмоциональность).
Тест "приподнятый крестообразный лабиринт" (ПКЛ) использовали в базовой модификации [3, 11], регистрируя в течение 5 мин: латентный период первого захода в рукав, время пребывания в открытых рукавах и на центральной площадке, число заходов в открытые и закрытые рукава, число выходов на центральную площадку.
Тест "условный рефлекс пассивного избегания" (УРПИ). Выработку условного рефлекса пассивного избегания у животных проводили в стандартной установке [2, 3]. Воспроизводимость УРПИ оценивали через 24 и 72 ч после обучения в течение трех минут, регистрируя латентный период первого захода в темный отсек, время нахождения в темном отсеке, число переходов между отсеками.
Полученные результаты обрабатывали статистически, используя t-критерий Стьюдента для сравнения независимых выборок. Для множественных сравнений использовали r-критерий Стьюдента с поправкой Бонферрони.
Результаты и их обсуждение
Контрольные мыши BALB/C и C57BL/6, помещенные в ОП, проявляли разные фенотипы поведения. Мыши C57BL/6 характеризовались высоким уровнем двигательной активности (горизонтальной и вертикальной), а также низким уровнем дефекаций. У мышей линии BALB/C в ОП наблюдалась реакция замирания с высоким уровнем дефекаций (рис. 1). Как показано ранее [9, 12] такие фенотипы поведения отражают высокий уровень исследовательской активности (мыши C57BL/6; "активный" фенотип ЭСР) и выраженную реакцию страха (мыши BALB/C; "пассивный" фенотип ЭСР).
Pис.1. Влияние мексидола (100мг/кг внутрь) на поведение mi,пней линий BALB/C (а) и C57BL/6 (б) в тесте "открытое поле" (М ± SEM).
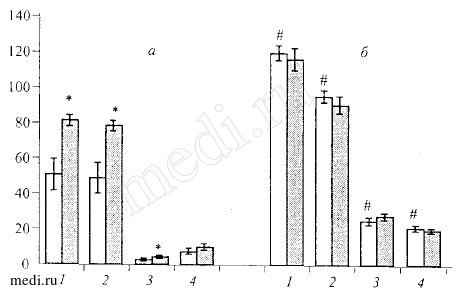
По оси абсцисс: 1 - общая двигательная активность, 2 - горизонтальная двигательная активность, 3 - вертикальная активность, 4 - исследовательская активность; по оси ординат: среднее число реакций животных зa 3 мин. Светлые столбики - контрольные животные, темные столбики животные, получавшие мексидол. * - р<0,05 по сравнению с контролем, # - р<0,05 между инбредными линиями.
Под влиянием мексидола (100 мг/кг) у мышей BALB/C наблюдалось увеличение горизонтальной двигательной и вертикальной активности, а также снижение числа фекальных болюсов (эмоциональности). Изменений в поведении "активных" мышей (C57BL/6) выявлено не было (рис. 1). Полученные результаты подтверждают полученные ранее данные, показавшие активацию поведения мышей BALB/C при использовании мексидола в более низкой дозе - 50 мг/кг [8]. У беспородных мышей мексидол в дозе 100 мг/кг не вызывал достоверного изменения исследуемых параметров.
Рис.2. Влияние мексидола (100 мг/кг внутрь) на латентный период первого захода в темный отсек (а) и число переходов между отсеками (б) в тесте "условный рефлекс пассивного избегания" у мышей инбредных линий и беспородных мышей (M+SEM).
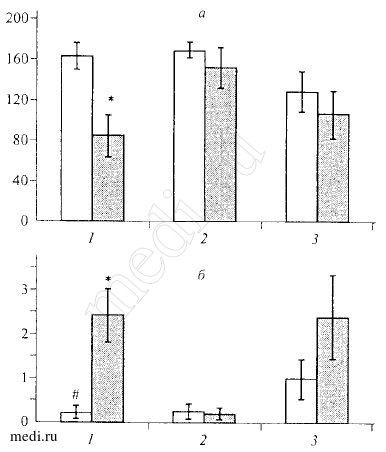
По оси абсцисс: 1 - мыши линии BALB/C, 2 - мыши линии C57BL/6, 3 - беспородные мыши; по оси ординат: а - время, с; б - число переходов. Светлые столбики - контрольные животные, темные столбики - животные, получавшие мексидол. * - р<0,05 по сравнению с контролем, # - р<0,05 по сравнению с беспородными животными.
Сравнительный анализ поведения мышей линий BALB/C и C57BL/6, проведенный в тесте ПКЛ, показал, что указанные линии животных практически не различаются по большинству оцениваемых в данном тесте показателей поведения. Только время задержки мышей BALB/C в центре лабиринта было достоверно выше аналогичного показателя, регистрируемого у животных линии C57BL/6 (таблица). Сходные данные получены другими авторами [6].
Таблица. Влияние мексидола (100 мг/кг, внутрь) на мышей инбредных линий и беспородных мышей в тесте "приподнятый крестообразный лабиринт" (М± SEM)
| Параметр теста | Мыши | |||||
| BALB/C | C57BL/6 | беспородные | ||||
| Контроль (n=15) |
Мексидол (n=17) |
Контроль (n=9) |
Мексидол (n=9) |
Контроль (n=17) |
Мексидол (n=17) |
|
| Время пребывания в открытых рукавах, с | 20,53 ± 2,55 | 39,76 ± 5,99* | 12,67 ± 3 | 17,33 ± 4,23 | 13,29 ± 2,45 | 13,71 ±2,9 |
| Время пребывания на центральной площадке, с | 49 ± 4,04 | 41,18 ± 4,56 | 28,78 ± 2,16# | 33,44 ± 3,46 | 45,76 ± 6,51 | 55,53 ± 8,28 |
| Латентный период первого захода в рукав, с | 2,6 ± 0,8 | 3,94 ± 0,52 | 2,33 ± 0,5 | 1,11 ±0,11* | 1,77 ± 0,33 | 1,94 ±0,29 |
| Число заходов в открытые рукава | 3,13 ± 0,39 | 3,47 ± 0,58 | 2,89 ± 0,59 | 3,67 ± 0,78 | 2 ± 0,4 | 2,29 ± 0,44 |
| Число заходов в закрытые рукава | 7,87 ± 0,7 | 8,77 ± 1,03 | 8,22 ± 1,1 | 8,11 ± 1,01 | 10,71 ± 0,99 | 9,12 ± 0,54 |
| Число выходов на центральную площадку | 6,53 ± 0,74 | 6,06 ± 0,7 | 6,33 ± 0,71 | 5,78 ± 0,57 | 7,41 ±0,91 | 7,65 ±0,71 |
| Общее число заходов в рукава | 11 ± 1,01 | 12,2 ± 41,34 | 14,11 ± 1,62 | 11,78 ± 1,66 | 12,71 ± 1,07 | 11,41 ± 0,63 |
# - р<0,05 между инбредными линиями.
Мексидол увеличивал время нахождения в открытых рукавах у мышей BALB/C (таблица), что указывает па его выраженное анксиолитическое действие. У мышей C57BL/6 под действием препарата уменьшался латентный период первого захода в рукав. На поведение беспородных мышей мексидол влияния не оказывал.
Поведение контрольных BALB/C и C57BL/6 в тесте УРПИ не различалось. Большую часть тестируемого времени животные проводили в светлом отсеке.
Мексидол изменял поведение животных в тесте УРПИ на третьи сутки после стресса. Препарат снижал латентный период первого захода мышей BALB/C в темный отсек (рис. 2, а) и в 3,6 раза увеличивал время их нахождения в опасном отсеке, повышая в то же время число переходов между отсеками (рис. 2,6). Изменений в поведении мышей C57BL/6 под влиянием мексидола не наблюдалось. Полученные результаты демонстрируют, что мексидол активирует поведение мышей BALB/C (уменьшение латентного периода первого захода в темный отсек и возросшее число переходов между отсеками) и не влияет на "активных" животных (C57BL/6), проявляя свойства селективного анксиолитика.
Таким образом, мексидол (100мг/кг) в условиях всех трех тестов устраняет реакцию страха, freezing, у мышей BALB/C, не оказывая при этом седативного действия на мышей C57BL/6, что свидетельствует о его селективном анксиолитическом действии на животных с "пассивным" фенотипом ЭСР.
Возможно, различие в антистрессорном действии мексидола у мышей BALB/C и C57BL/6 обусловлено межлинейными различиями в бензодиазепиновой рецепции у лабораторных животных, отличающихся по фенотипу поведения в стрессовых ситуациях. Из литературы известно, что после эмоционально- стрессового воздействия в тесте ОП наблюдается резкое падение связывания меченного лиганда (3Н-диазепама) у мышей BALB/C, в то время как у мышей C57BL/6 параметры рецепции не меняются [10]. Уменьшение бензодиазепиновой рецепции у мышей BALB/C приводит к ослаблению у них ГАМК-ергических процессов, что имеет следствием анксиогенный эффект, сопровождаемый реакцией замирания. У мышей C57BL/6 подобные изменения не возникают, что и обусловливает их стрессустойчивость [10]. Анксиоселективный препарат афобазол препятствует развитию нарушений в ГАМК-бензодиазепиновом комплексе у мышей BALB/C ("пассивных" животных) при стрессе и проявляет избирательный анксиолитический эффект [9]. Можно предположить, что подобный механизм действия имеет и мексидол. Не обладая прямым аффинитетом к бензодиазепиновым и ГАМК-рецепторам, мексидол, вероятно, может оказывать на них модифицирующее действие, аллостерически потенцируя рецептор лиганда и активируя ионные каналы, тем самым усиливая способность рецепторного комплекса к связыванию [8, 13].
Выводы
- Мексидол увеличивает у мышей BALB/C время нахождения в открытых рукавах и не изменяет поведение мышей C57BL/6 в тесте "приподнятый крестообразный лабиринт".
- Мексидол уменьшает латентный период первого захода в темный отсек и повышает число переходов между отсеками у мышей BALB/C в тесте "условный рефлекс пассивного избегания", что свидетельствует о его анксиолитическом эффекте.
- Селективное анксиолитическое действие мексидола проявляется у мышей BALB/C с "пассивным" фенотипом эмоционально-стрессовой реакции в условиях "избегаемого" и "неизбегаемого" эмоционального стресса.
ЛИТЕРАТУРА
- Ю. А. Бледное, М. М. Козловская. М. Маула, Тез. докл. на 2-ой Международ, конференции: "Выживание человека: резервные возможности и нетрадиционная медицина", Москва, 24 - 25 августа, 1994, Москва (1994), сс. 61 - 62.
- Я. Буреш, О. Бурешова, Дж. П. Хьюстон, Методики и основные эксперименты по изучению мозга и поведения, Высшая школа, Москва (1991).
- Т. А. Воронина, С. Б. Середенин, Методические указания по изучению транквилизирующего (анксиолитического) действия фармакологических веществ /Руководство по экспериментальному (доклиническому) изучению новых фармакологических веществ, Ремедиум, Москва (2000), сс. 26- 130.
- Т. А. Воронина, Психофармакол. и биол. наркол., № 1,2- 12 (2001).
- К. М. Дюмаев, Т. А. Воронина, Л. Д. Смирнов, Антиоксиданты в профилактике и терапии патологий ЦНС, Изд. Инст-та биомед. химии РАМН, Москва (1995).
- Мона Ассад Маула, Автореф. дис. канд. биол. наук, Москва (1996).
- Г. Г. Незнамов, Е. С. Телешова, Т. П. Сафарова и др., Материалы международного симпозиума "Биоантиоксидант ", Тюмень (1997), сс. 85 -87.
- С. Б. Середенин, Ю. А. Бледнов, М. JI. Гордей и др., Хим.-фарм. ж., № 2, 134- 137 (1987).
- С. Б. Середенин, Т. А. Воронина, Г. Г. Незнамов и др., Вестн. РАМН, № 11, 3 - 9 (1998).
- С. Б. Середенин, М. А. Яркова, М.В.Воронин, Экспер. и клин, фармакол., 64(1), 63 - 65 (2001).
- Е. Pellow, P. Chopin, S. Е. File, and М. Briley, J. Neurosci. Methods., 14, 149- 167 (1985).
- S. B. Seredenin and Y. A. Blednov, Biological basis of individual sensitivity to psychotropic drug, Edinburgh (1995), 25 - 38.
- Т. A. Voronina and S. В. Seredenin, Ann. 1st. Super. Sanita., 24, 461 - 466 (1988).



