 polysan.ru
polysan.ru
Применение Цитофлавина PRO способствует:
- увеличению устойчивости к стрессам;
- повышению умственной и физической работоспособности
Взаимоотношения тревоги и сна в эксперименте и клинике
СтатьиОпубликовано в журнале:
Журнал неврологии и психиатрии, 4, 2017; Вып. 2
https: //doi.org/10.17116/jnevro20171174212-18
Е.В. Вербицкий
ФГБУ «Институт аридных зон» Южного научного центра РАН, Ростов-на-Дону, Россия
Высокая тревожность — черта личности, во многом детерминированная генетически, которая обусловлена функционированием лимбической системы и деятельностью других церебральных образований, участвующих в поддержании бодрствования и развитии сна. Анализ экспериментов на животных и клинических наблюдений указывает на то, что эта черта, определяя особое реагирование индивида, способствует развитию тревоги и тревожных расстройств, что за счет угнетения ГАМК-бензодиазепиновых тормозных влияний негативно сказывается на продолжительности и качестве ночного сна. Феназепам за счет анксиолитических свойств дезактивирует очаги возбуждения в нервной системе, купируя негативные проявления тревоги, что оказывает сомногенное действие, нормализующее нарушения сна.
Ключевые слова: тревога, тревожность, сон, качество сна, феназепам, бензодиазепиновые рецепторы, анксиолитики.
Anxiety and sleep in experiment and clinic
E.V. Verbitsky
Institute of Arid Zones of the Southern Scientific Centre of Russian Academy of Sciences, Rostov-on-Don, Russia
High anxiety is a genetically determined personal trait. It is produced by limbic system and other cerebral structures involved in maintenance of wakefulness and development of sleep. Analysis of experiments on the animals and clinic observations showed that animals and individuals with high personal anxiety have a high risk of anxiety disorders. Inhibition of GABA-benzodiazepine system which is typical for high anxiety dramatically influences sleep duration and sleep quality. Phenazepamum due to its anxiolytic properties deactivates the excitation focus in the brain, jugulates anxiety and normalize sleep.
Keywords: anxiety, personal trait anxiety, sleep, sleep quality, phenazepamum, benzodiazepine receptors, anxiolytics, insomnia.
За последние десятилетия на значительной части земного шара происходили межэтнические или межнациональные конфликты, политические обострения, назревали экономические кризисы, возникали военные противостояния. Все это способствовало усилению социальных и других противоречий в обществе и так или иначе отразилось на каждой отдельной личности, которую прежде не настолько тревожили проблемы социума и экономики, кризисные ситуаций и катастрофические события. Не случайно во всех странах мира наблюдается увеличение заболеваний, связанных с высокой тревожностью и патологической тревогой. Мы становимся свидетелями появления ранее не известных заболеваний: фибромиалгии, синдрома хронической усталости и др. В то же время такие заболевания, как гипертония, ожирение, неспецифические иммунодефицитные нарушения, изменили свои параметры и стали чаще встречаться в более «злокачественных» формах [1, 2]. Таким образом, пришло время рассматривать указанные события в единой связи с расширением пределов неадекватного реагирования, когда граница, отделяющая высокую тревожность от тревожных расстройств, слабо различима, что способствует нарушению адекватного бодрствования и негативно сказывается на развитии ночного сна.
Подходы к изучению тревожного реагирования
Изучение тревожного реагирования имеет свою историю, в которой российские ученые сыграли достойную роль. Физиологические и клинические варианты страха, тревожности, тревоги, расстройств сна животных и человека со времен павловской школы неврозов детально описаны в российской, а также в западной научной литературе прошлого века [3—5]. Медицина сна подтверждает взаимосвязь некоторых сомнологических нарушений и особенностей тревожного реагирования. В США, например, согласно данным Национального института здоровья, сон нарушен у 30—40% городского населения, причем более чем в 60% случаев прослеживается связь нарушения сна с нарастанием тревожного реагирования (от хронических страхов до панических атак). Поэтому нарушения сна входят в список первичных симптомов таких заболеваний, как депрессия, шизофрения, аутистические синдромы, и в ряде случаев считаются индикаторами приближающей декомпенсации [2, 6]. Таким образом, становится понятен интерес врачей к моделированию патологической реорганизации тревожного реагирования и его связи со сном в экспериментах на животных.
Что же является общим для животных и человека, когда они попадают в новую обстановку? Прежде всего, это активация нервной системы и запуск ориентировочного реагирования, необходимого для уточнения потенциальной опасности, которая может повышать или снижать ориентировочное реагирование и сказываться на развитии сна.
Сначала в изучении тревожности как склонности к тревожным расстройствам использовались модели с интенсивными раздражителями. Животных подвергали воздействию электрического тока, иммобилизации и др. Сейчас не вызывает сомнения то, что эти подходы чаще провоцировали стресс, а не тревожность [7]. Позднее наметились другие тенденции и особую популярность приобрели тесты, более близкие к реальности. В частности, анализировали поведение животных в новых условиях: в открытом поле, в норковой камере, в крестообразном приподнятом лабиринте, в соседстве с агрессивной особью [2].
Другое перспективное направление изучения тревожности, которое тоже возникло в эпоху снижения интенсивностей стимулов, — исследование проявлений тревожности у приматов, хищников, грызунов по проявлению атипичной, нецеленаправленной или смещенной активности [8]. Здесь внимание уделялось мелкой двигательной активности и особенно различным проявлениям аномального груминга — гигиенической процедуры животных, например вылизывания, имеющей социальное значение. Позже пришло понимание того, что груминг обладает существенной гетерогенностью, к тому же выяснилось, что не для всех животных и не во всех ситуациях он является маркером тревожности. Выяснилось, что сложная структура груминга довольно часто проявляется только во взаимосвязи с реализацией других форм поведения или выполнения физиологических функций. Решение таких задач требует секвенциального подхода к анализу физиологических и поведенческих отражений тревожности [8, 9].
По классическим представлениям Ч. Спилбергера, тревожность имеет личностные (базовые) и ситуативные (реактивные) компоненты [10, 11]. Однако со временем стало понятно, что высокая базовая тревожность неизбежно провоцирует проявление ситуативной тревожности [12, 13]. Поэтому модели для изучения ситуативной тревожности, не учитывающие уровень базовой тревожности, не вполне адекватны на современном этапе понимания природы тревожности и ее границ с тревогой (тревожными расстройствами) [3]. Важны модели, позволяющие изучать базовую тревожность на животных в естественных условиях, в сочетании ее проявлений с другими физиологическими функциями организма. Уместно вспомнить подход, забытый на многие годы, который свидетельствует о том, что поведение животного, вынужденного исследовать новизну ситуации, может существенно отличаться от естественного, спонтанного поведения, когда животное само решает, исследовать ситуацию сейчас или в этом нет насущной необходимости [13]. Развитие этого подхода позволило установить, что принуждение животного к оцениванию новизны ситуации неизбежно повышает ситуативную тревожность. В результате были разработаны новые направления изучения тревожности, учитывающие не отдельные маркеры, а систему признаков [2, 14].
Физиология тревожности и действие анксиолитиков
С нейрофизиологических позиций все варианты высокой тревожности реализуются при тесном участии образований паралимбического комплекса. Лимбическая система состоит из филогенетически старых отделов переднего мозга, которые образуют корковую область этой системы, и их производных, которые объединяются в группу подкорковых образований. К корковым областям относят: гиппокамп, парагиппокампальную извилину, поясную извилину, а также обонятельные луковицы и бугорки, орбитофронтальную, островковую и частично височную кору. В число подкорковых структур включают: миндалину, септальные ядра, переднее ядро таламуса, а также преоптическую область, гипоталамус, мамиллярные тела и другие образования [15—17] (см. рисунок).
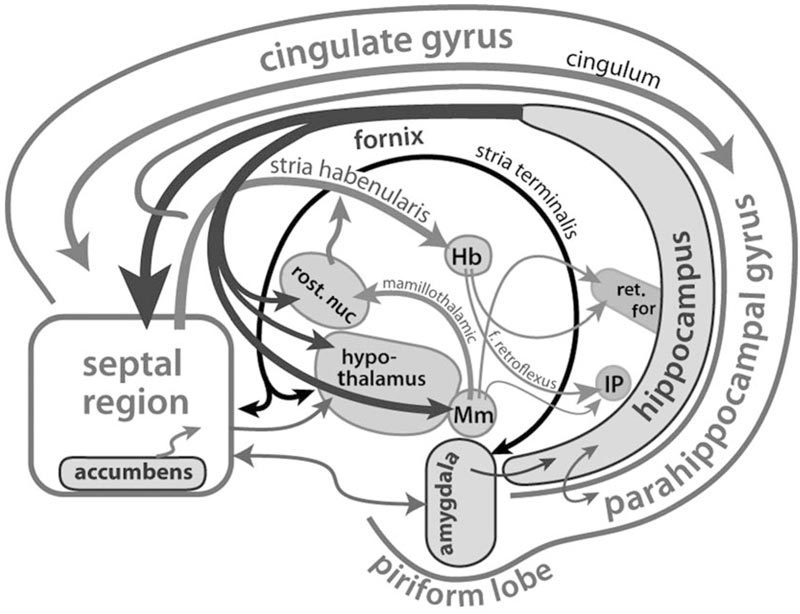
Рисунок. Образования лимбической системы [34].
Accumbens — септальный комплекс; amigdala — миндалевидный комплекс; cingulate gyrus — область поясной извилины; IP — интерпедункулярный комплекс; fornix — свод; Hb — хабенулярный комплекс; hippocampus — гиппокамп; hypothalamus — гипоталамус; Mm — мамиллярный комплекс; parahippocampal gyrus — область парагиппокампальной извилины; piriform lobe — область височной коры; ret. for — образования ретикулярной формации; rost. nuc — передние ядра неспецифического таламуса; septal region — область перегородки.
Афферентные и эфферентные связи образований лимбической системы достаточно сложны и разнообразны. Наиболее мощные связи обнаружены между лимбической системой и гипоталамусом. Так, гипоталамус и мамиллярные тела соединяются с гиппокампом и септальной областью посредством пучка волокон свода. Гипоталамус связан с миндалиной посредством терминальной полоски и амигдалофугального пучка волокон, а с фронтальными частями обонятельного мозга — с помощью медиального пучка переднего мозга. Со средним и промежуточным мозгом, а также с расположенными ниже образованиями мезенцефалической и находящимися выше структурами таламической ретикулярной формации лимбическая система взаимодействует через гипоталамус, мамиллярные тела и другие образования головного мозга [17, 18].
Поскольку для лимбической системы характерны многочисленные замкнутые цепи, по которым может циркулировать возбуждение, считается, что образования этой системы отвечают не только за дифференциацию новизны и формирование эмоционального поведения, но также участвуют в формировании следов памяти [5, 19]. Поэтому лимбикоретикулярной системе отводится ключевая роль в приспособительном поведении к изменяющимся условиям внешней или внутренней среды организма [20]. Судя по всему, в лимбико-ретикулярной системе поступающая информация сопоставляется с опытом в памяти и запускает те поведенческие реакции, которые наиболее адекватны ситуации, определяя характер тревожного реагирования.
В течение нескольких десятилетий было много сделано для уточнения представлений о том, как сигналы от гипоталамуса, мамиллярных тел и лимбических областей среднего мозга поступают к поясной извилине через переднее ядро таламуса и другие образования, а затем к лобным областям коры при участии медиального и медиодорсального таламических ядер. Это позволяет детализировать представления об окружающей среде, отделив биологически и социально значимые признаки (из височных областей коры), а также уточнить сведения о внутренней среде (из образований гипоталамуса) в ходе конвергации их сигналов на нейронах миндалины. Надо полагать, что это и есть тот нейрофизиологический субстрат, который необходим для образования прочных связей в памяти, объединяющих сигналы внешней среды с состоянием внутренней среды организма для адаптивного реагирования [12, 13, 20].
В последнее время, помимо описанных выше образований головного мозга, с уровнем тревожности связывают деятельность коры больших полушарий, в частности таких ее областей, как ее теменные регионы, а также образований медиального таламуса, базальных ганглиев и хвостатого ядра. Однако пока нет единства мнений по вкладу этих структур в уровни тревожности. Следует отметить, что направленность перестроек указанных образований при возрастании уровня тревожности, когда активируются arousal-реакции (реакция ЭЭГ-активации), указывает на связь тревожного реагирования с механизмами переключения от сна к бодрствованию. В пользу этого свидетельствуют данные о снижении тревожности после сеансов угнетения активности ядерных образований медиального таламуса [15]. С другой стороны, ГАМК-тормозную систему, играющую важную роль в снижении бодрствования и углублении медленноволнового сна, также связывают с уровнем тревожности.
Однако связь тревожности с уровнем активации нервной системы не отличается линейностью. Еще в 1995 г. был обнаружен феномен анксиоличности слабых доз некоторых ГАМК-литических препаратов (коразол, пенициллин), которые до этого были известны как классические анксиогены. Предположительно, умеренное возбуждение нервной системы провоцирует эмоционально-положительные состояния (pleasurable emotioms), которые приводят к неожиданному снижению тревожности до тех пор, пока растущая активация мозга снова не начнет провоцировать рост тревожности [13]. Общность тревожных и депрессивных механизмов часто объясняют участием ГАМК-бензодиазепиновой тормозной системы, деятельность которой затрагивает более 70% нейронов мозга, особенно ионотропных ГАМКА- и метаботропных ГАМББ-рецепторов. Оба типа рецепторов вовлечены в регуляцию интегративных процессов мозга, таких как сон, память, эмоции, тревожность, эпилепсия. В настоящее время известно, что гены, кодирующие субъединицы ГАМКА-рецепторов, локализованы в виде пяти кластеров на разных хромосомах. Два гена, кодирующие ГАМКБ, Б1- и Б2-единицы, расположены на двух других хромосомах. По данным мутагенеза, гены, кодирующие 7 единиц ГАМКА-рецепторов, связываются с выраженностью тревожности [3, 18]. Так полиморфизм ГАМКА-ергических генов человека связывается с тревожными расстройствами [2, 21].
Природу анксиолитических эффектов целесообразно рассмотреть на примере феназепама — производного бензодиазепина (7-бром-5-(ортохромфенил)-2-3-дигидро-1Н-1,4-бензодиазепин-2-ОН). Он, являясь бензодиазепином, действует как аллостерический модулятор ГАМКА-рецепторов в мамилярных телах заднего гипоталамуса лимбической системы головного мозга, которые участвуют в формировании тревожно-эмоциональной реактивности организма [22]. Там феназепам связывается с γ2или с α1- (гипнотический эффект), α2- (анксиолитический эффект), α3-, α5-субъединицами рецепторов. Учитывая то, что ГАМКА-рецепторы широко представлены во всех структурах головного мозга, определить преимущественную область влияния препарата довольно сложно.
В настоящее время очевидно, что действие феназепама на ГАМКА-рецепторы в вентролатеральном преоптическом ядре гипоталамуса запускает сон, облегчая при этом ингибиторную ГАМКергическую передачу в целом. Под его влиянием усиливается ГАМК-торможение нейронов голубого пятна и падает высвобождение норадреналина, особенно в области префронтальной коры. В это время на ЭЭГ регистрируются гигантские дельта-волны, генерирование которых частично подавляет развитие быстрого сна. При этом наблюдается активация ГАМКергических нейронов вентролатерального преоптического ядра гипоталамуса, судя по росту экспрессии сигнальных молекул c-Fos, и регистрируется торможение большинства корковых нейронов, мембраны которых гиперполяризованы и подвержены обширным синхронным колебаниям клеточных потенциалов в ритме дельта-волн. Следует отметить, что мутации в α1-субъединице ГАМКА-рецепторов ослабляют гипногенный эффект бензодиазепинов, а мутации в α2-субъединице блокируют их анксиолитическое влияние [21, 23].
Прием феназепама, как и других бензодиазепинов, в ряде случаев может приводить к развитию зависимости [24, 25] при длительном приеме в больших дозах (более 4 мг/сут). Во избежание этого были разработаны препараты, преимущественно влияющие на деятельность α1-субъединицы ГАМКА-рецепторов, так называемые Z-препараты. Селективность воздействия на рецепторы лишает эти препараты клинически выраженного противотревожного эффекта. Они отличаются между собой продолжительностью весьма короткого действия и не лишены нежелательных последствий. Природа таких последствий обусловлена значительной представленностью α1-субъ единиц ГАМКА-рецепторов в обширных кортикальных областях и в глубоких образованиях головного мозга, которые отличаются: архитектурой связей, медиаторной природой межнейронных взаимодействий, что в цикле сон—бодрствование проявляется масштабной реорганизацией нейрональной активности. Это подтверждается тем, что полиморфизм гена Б1-субъединицы ГАМКБ-рецепторов недавно был ассоциирован с паническими атаками, а локус 5g34 хромосомы 5, содержащий кластер генов субъединиц ГАМКА, — с расстройствами настроения и простыми по структуре тревожными расстройствами. Кроме того, получены данные о связи уровня тревожности и тревожных расстройств с обменом глутаматдекарбоксилазы, ГАМК-трансаминазы и других соединений, активность которых обусловлена экспрессией гена GAD65 [21].
Связь между тревожностью и развитием сна
Тормозное состояние в медленноволновом сне наступает за счет переднего гипоталамуса, а также неспецифического таламуса и части черного вещества, образующих импульсы, которые приводят к гиперполяризации мембран большинства корковых нейронов и синхронным колебаниям постсинаптических потенциалов на обширных регионах коры больших полушарий. Основными медиаторами медленного сна являются ГАМК и серотонин, а также аденозин и простагландин D2 в древней тормозной системе оболочек мозга и хорального сплетения [17, 18]. Медленный сон издавна связывали с репаративными функциями, накоплением макроергических связей. Начало и углубление медленного сна контролируется нейронами вентролатеральной и срединной преоптической области переднего гипоталамуса. Важную роль в поддержании медленного сна играет также парафасцикулярная зона рострального отдела продолговатого мозга, которая угнетает деятельность активирующих глутаматергических нейронов покрышки моста. А формирование самой продолжительной стадии медленного сна — стадии сонных веретен — выполняют ретикулярное ядро, срединный центр и другие образования неспецифического таламуса [26].
Развитие быстрого сна отличается от медленного формированием активированного состояния с подавлением моторной активности, чему способствуют образования ретикулярной формации продолговатого мозга и варолиева моста, где трансмиттерами являются ацетилхолин, глутамат и аспаргиловая кислота [27]. Во время быстрого сна наблюдается сновидческая активность, которую связывают с «эмоциональной разрядкой» мотиваций, не реализованных в бодрствовании, а также с поддержанием поисковой активности. Депривация быстрого сна у экспериментальных животных, которые размещались на маленьком островке и при снижении мышечного тонуса падали в воду, приводила к увеличению проявлений тревожности, повышению возбудимости, агрессивности, двигательного беспокойства и в целом меняла характер ориентировочного реагирования [2]. Предполагают, что причиной этого является тесная взаимосвязь дорсальных и медианных ядер шва посредством серотонинергических волокон с миндалиной. Также считают, что повышение содержания серотонина в миндалине способствует появлению быстрого сна, а его снижение влияет на глубину сна и переходы к бодрствованию при тесном участии других образований переднего мозга. Причем выключение гена, контролирующего развитие рецепторов серотонина, характеризуется пониженной активностью и боязнью открытого пространства, что характерно для высокого уровня тревожности [21]. Угнетение рецепторов серотонина в моделях на животных приводило к аномалиям в соматосенсорной коре, нарушениям медленноволнового сна, гипоактивности, высокой тревожности, тахикардии, гипертензии и извращению чувствительности к психотропным препаратам. Управление запуском быстрого сна происходит в ГАМКергическом центре вентральной области продолговатого мозга. Здесь инициируется «торможение торможения» для активации нейронов гиппокампа, миндалины, отдельных зон коры мозга при сохранении минимального тонуса мускулатуры, что исключает движение [17].
Ранее было установлено, что у обследуемых, которых по современным оценкам [10, 13] можно классифицировать как индивидов с высоким уровнем тревожности, фаза быстрого сна длилась дольше, да и потребность во сне у них была выше [27—29]. При этом они были более чувствительны к насущным проблемам, им было свойственно принимать все близко к сердцу и реагировать на рост трудностей или появление заболеваний увеличением продолжительности быстрого сна [20, 30]. Чаще всего самопроизвольное сокращение времени сна приходилось на периоды относительного благополучия индивида, когда он с интересом работал, получал удовлетворение от прожитого и когда проявления его ситуативной тревожности были невысокими. Если же количество неразрешимых проблем в бодрствовании возрастало, снижалось настроение, росли проявления тревожности, то увеличивалась общая потребность организма во сне, особенно в фазе быстрого сна.
В настоящее время показано, что индивиды с высоким уровнем личностной тревожности имеют характерные особенности в ЭЭГ бодрствования. В отличие от обследуемых с низким уровнем тревожности им свойственны: меньший фронтоокципитальный градиент амплитуды, в спектре их ЭЭГ часто фиксируются высокочастотные компоненты и спонтанные активации [27, 31]. Меньшая амплитуда отличает у них средне- и длиннолатентные компоненты вызванных потенциалов на слуховые и зрительные стимулы, габитуация которых при увеличении количества раздражителей существенно снижена за счет, вероятно, угнетения синхронизирующих влияний со стороны таламокортикальной неспецифической системы. Однако наиболее существенные отличия наблюдают во время сна у обследуемых с высоким уровнем тревожности: сужение глубины сна, увеличение неглубокого сна за счет редукции глубокого медленного сна. Кроме того, сну таких больных свойственна большая сегментация стадий сна со значительным количеством микроактиваций нервной системы, а также с множеством эпизодов ЭЭГ-бодрствования среди сна [29]. Оценки спектральной плотности ЭЭГ у них более плоские в спокойном бодрствовании, а также во время ночного сна по сравнению с обследуемыми, которым свойствен или был навязан фармакологически низкий уровень тревожности. Эти факты подтверждают, что нейрофизиологические механизмы тревожности тесно связаны с активационными механизмами мозга [31, 32].
В связи с этим интересно проследить динамику нормализации сна под влиянием такого гипнотика, как феназепам в дозе 2 мг в сутки, у пациентов с разными формами тревожных расстройств. Так, общими для всех пациентов были отчетливые анксиолитические и гипногенные эффекты, которые проявлялись в редукции проявлений тревоги и улучшении ночного сна на 3—7-й день терапии. Однако у больных с простыми по структуре формами тревожных расстройств, которые характеризовались чувственными, образными эмоциональными фобиями, непосредственно связанными с тревогой, нарушения засыпания снижались на 87% на 14-й день терапии. Нарушения ночного сна снижались практически до нуля уже на 3-й день. Несколько менее выражено уменьшались нарушения пробуждения — на 30% к 14-му дню терапии. Действие препарата, как правило, не сопровождалось утренней сонливостью и вялостью [33].
У пациентов с тревожными расстройствами, объединяющими сенесто-ипохондрические и идеообсессивные расстройства типа навязчивой ипохондрии, действие феназепама в дозе 2 мг в сутки проявлялось в уменьшении выраженности тревоги, раздражительности и аффективной неустойчивости, а также чувственно окрашенных фобий. Что же касается сомнологических нарушений, то динамика эффектов препарата была несколько иной. Так, в частности, нарушения засыпания и ночного сна у таких больных уменьшались на 67% на 14-й день приема препарата.
Иные закономерности в динамике сомнологических показателей были выявлены у пациентов со сложной структурой тревожных расстройств. Особенности влияния феназепама у этих больных в дозе 2 мг в сутки проявлялись в выраженной редукции тревоги и в снижении сомнологических нарушений. В частности, происходило уменьшение нарушения засыпания на 85% при курсовом приеме препарата 14 дней. Нарушения ночного сна у этих больных падали до нуля уже на 7-й день проведения курсовой терапии. А нарушения пробуждения снижались на 72% к 14-му дню терапии [33].
Таким образом, эффективность курсовой терапии феназепамом зависела от структурной композиции тревожных расстройств. В наиболее полной мере клинико-фармакологические эффекты препарата реализовались у пациентов с простыми по структуре тревожно-астеническими, тревожными и тревожно-фобическими расстройствами в тех случаях, когда фобии демонстрировали тесную коморбидность с тревогой, а также отличались образностью и эмоциональной насыщенностью [33].
Сходные результаты были получены при изучении сна животных, дифференцированных на группы особей с высоким и низким уровнем базовой тревожности. Оказалось, что у особей с высокой тревожностью тоже наблюдается уменьшение времени стадий глубокого сна за счет роста неглубокого медленного сна на фоне значительного увеличения сегментаций. Пороги на акустические раздражители у этих животных были ниже в спокойном бодрствовании, а также в медленном сне. А габитуация вызванных потенциалов была меньше у особей с высокой тревожностью, чем у особей с низким уровнем тревожности, что имело сходство с закономерностями габитуации вызванных ответов у обследованных добровольцев без нарушений здоровья, дифференцированных на индивидов с высокой и низкой личностной тревожностью [2, 10].
Заключение
Таким образом, в настоящее время не вызывает сомнения связь уровня тревожности в бодрствовании с развитием сна. Судя по всему, такая связь обусловлена взаимодействием образований кортикостриопаллидарной и лимбической систем мозга, где особая роль отводится ГАМК-ергическим тормозным влияниям. Вне всякого сомнения, высокий уровень тревожности во многом определяет характер тревожного реагирования и индивидуальные особенности мотивационно-эмоционального поведения. На примере приема типичного анксиолитика (феназепам) показано снижение тревоги и нормализация сна, выраженные лучше у пациентов с простой структурой тревожных расстройств. Важный вклад в эти процессы, судя по экспериментам на животных, вносят медиобазальные участки коры головного мозга, передняя область гипоталамуса и древняя аденозиновая система, которые наряду с другими образованиями переднего мозга участвуют в обеспечении бодрствования и поддержании сна. Главная роль отводится задней области гипоталамуса и ядрам миндалины, которые определяют уровень тревожности в бодрствовании и вносят свой вклад в развитие быстрого сна. Несмотря на очевидность связи механизмов тревожности и тревожных расстройств с деятельностью активационных образований головного мозга, эти процессы требуют дальнейшего исследования.
Статья подготовлена при поддержке фармацевтической компании ПАО «Валента Фарм».
Литература/References
- Golbin A, Kraviyz H, Keith L. Sleep psychiatry. London—New York: Taylor & Francis; 2004. Golbin A, Kraviyz H, Keith L. Sleep psychiatry. London— New York: Taylor & Francis; 2004.
- Вербицкий Е.В. Сон и тревожность. Ростов-на-Дону: Изд. Южного научного центра РАН; 2008. [Verbitskii EV. Son i trevozhnost’. Rostov-naDonu: Izd. Yuzhnogo nauchnogo tsentra RAN; 2008. (In Russ.)].
- Александровский Ю.А. Пограничные психические расстройства. М.: Медицина; 1993. [Aleksandrovskii YuA. Pogranichnye psikhicheskie rasstroistva. M.: Meditsina; 1993. (In Russ.)].
- Eysenck MW. Anxiety and attention. Anx Res. 1988;1:9-15.
- Симонов П.В. Лекции о работе головного мозга. Потребностно-информационная теория высшей нервной деятельности. М.: Изд. Инс. психол. РАН; 1998. [Simonov PV. Lektsii o rabote golovnogo mozga. Potrebnostno-informatsionnaya teoriya vysshei nervnoi deyatel’nosti. M.: Izd. Ins. psikhol. RAN; 1998. (In Russ.)].
- Бузунов Р.В. Бессонница в практике терапевта. Учебное пособие для врачей; 2009. sleepnet.ru/vracham/library/bessonnitsa-vpraktiket… [Buzunov RV. Bessonnitsa v praktike terapevta. Uchebnoe posobie dlya vrachei; 2009. sleepnet.ru/vracham/library/bessonnitsa-v-praktike…
- Калуев А.В. Стресс, тревожность и поведение: актуальные проблемы моделирования тревожного поведения у животных. Киев: Изд. КГУ; 1998. [Kaluev AV. Stress, trevozhnost’ i povedenie: aktual’nye problemy modelirovaniya trevozhnogo povedeniya u zhivotnykh. Kiev: Izd. KGU; 1998. (In Russ.)].
- Калуев А.В. Груминг и стресс. М.: Изд. АВИКС; 2002. [Kaluev AV. Gruming i stress. M.: Izd. AVIKS; 2002. (In Russ.)].
- Дерягина М.А., Бутовская М.Л. Этология приматов. М.: Изд. МГУ, 1992. [Deryagina MA, Butovskaya ML. Etologiya primatov. M.: Izd. MGU; 1992. (In Russ.)].
- Ханин Ю.Л. Краткое руководство к применению шкал реактивной тревоги и личностной тревожности. Л.: Изд. ЛНИИТЕК; 1976. [Khanin YuL. Kratkoe rukovodstvo k primeneniyu shkal reaktivnoi trevogi i lichnostnoi trevozhnosti. L.: Izd. LNIITEK; 1976. (In Russ.)].
- Spilberger CD, Gorsuch RL, Lushene RE. STAI manual for the state-trial anxiety inventory. New-York: Consulting Psychology Press Palo Alto; 1970.
- Хекхаузен Х. Мотивация и деятельность. В 2 т. Пер. с нем. под ред. Величковского Б.М. Т2. М.: Мир; 1986. [Khekkhauzen Kh. Motivatsiya i deyatel’nost. V 2-kh t. Per. s nem. pod red. Velichkovskogo BM. 2. M.: Mir; 1986. (In Russ.)].
- Вербицкий Е.В. Психофизиология тревожности. Ростов-на-Дону: Изд. Ростовского госуниверситета; 2003. [Verbitskii EV. Psikhofiziologiya trevozhnosti. Rostov-na-Donu: Izd. Rostovskogo gosuniversiteta; 2003. (In Russ.)].
- Kalueff A, LaPorte J. Experimental Animal Models in Neurobehavioral Research. Hardcover. New York: NOVA Science; 2009.
- Hartmann EL. The functions sleep. New Haven and London: Yale University Press; 1973.
- Ковальзон В.М. Основы сомнологии. М.: Изд. БИНОМ; 2011. [Koval’zon VM. Osnovy somnologii. M.: Izd. BINOM; 2011. (In Russ.)].
- Ковальзон В.М., Долгих В.В. Регуляция цикла бодрствование—сон. Неврологический журнал. 2016;6:316-322. [Koval’zon VM, Dolgikh VV. Regulyatsiya tsikla bodrstvovanie—son. Nevrologicheskii zhurnal. 2016;6:316-322]. https: //doi.org/10.18821/1560-9545-2016-21-6-
- Kovalzon VM. Ascending reticular activating system of the brain. Translational Neuroscience and Clinics. 2016;2(4):275-285. https: //doi.org/10.18679/CN11-6030/R.2016.034
- Купалов П.С. Механизмы замыкания временной связи в норме и патологии. М.: Медицина; 1978. [Kupalov PS. Mekhanizmy zamykaniya vremennoi svyazi v norme i patologii. M.: Meditsina; 1978. (In Russ.)].
- Borbely A, Hayaishi O, Sejnowski T, Altman J. The regulation of sleep. Human Frontier Science Programm: Strasbourg; 2000.
- Петров А.М., Гиниатуллин А.Р. Нейробиология сна: современный взгляд. Учебное пособие. Казань: КГМУ; 2012. [Petrov AM, Giniatullin AR. Neirobiologiya sna: sovremennyi vzglyad: Uchebnoe posobie. Kazan’: KGMU; 2012. (In Russ.)].
- Benzodiazepines: Good Practice Guidelines for Clinicians. All Ireland Public Health Repository. Department of Health (Ireland): 2002. (Last Updated: 2016-12-21). 28Р. https: //doi.org/10.14655/771574-773355
- Кузнецов С.Л., Афанасьев М.А. Значение гена раннего реагирования c-fos и продуктов его экспрессии в нейронах при различных воздействиях. Биомедицина. 2013;1(1):109-116. [Kuznetsov SL, Afanas’ev MA. Znachenie gena rannego reagirovaniya c-fos i produktov ego ekspressii v neironakh pri razlichnykh vozdeistviyakh. Biomeditsina. 2013;1(1):109-116. (In Russ.)].
- Левин Я.И., Полуэктов М.Г. Сомнология и медицина сна. Избранные лекции. М.: Медфорум; 2013. [Levin YaI, Poluektov MG. Somnology and Sleep Medicine. Selected Lectures. Somnologiya i meditsina sna. Izbrannye lektsii. M.: Medforum; 2013. (In Russ.)].
- Полуэктов М.Г. Сомнология и медицина сна. Национальное руководство памяти А.М. Вейна и Я.И. Левина. М.: Медфорум; 2016. [Poluektov MG. Somnology and Sleep Medicine. National Manual in Memory of A.M. Vein and Y.I. Levin. Somnologiya i meditsina sna. Natsional’noe rukovodstvo pamyati A.M. Veyna i Ya.I. Levina. M.: Medforum; 2016. (In Russ.)].
- Kryger MH, Roth T, Dement WC. Principles and Practice of Sleep Medicine, 6th ed. Philadelphia, PA, U.S.A: Elsevier; 2016.
- Сысоева Ю.Ю., Вербицкий Е.В. Индивидуальные циклические изменения дельта-активности мозга в медленном сне человека. Доклады Академии наук. 2012;444(1):1-5. [Sysoeva YuYu, Verbitskii EV. Individual’nye tsiklicheskie izmeneniya del’ta-aktivnosti mozga v medlennom sne cheloveka. Doklady akademii nauk. 2012;444(1):1-5. (In Russ.)].
- Sysoeva Yu, Verbitsky E. The Combination of High Trait Anxiety and Female Sex is Vulnerability Factor for Sleep Disturbances. Trait and State Anxiety: Assessment, Preductor and Outcomes. Nova Publishers, New York, Library of Congress Control Number 2016933393. ISBN: 978-163484-860-2 (eBook) 2016. novapublishers.com/catalog/product_info.php
- Сысоева Ю.Ю., Вербицкий Е.В. Характер активаций мозга во время ночного сна связан с личностной тревожностью индивидуума. Доклады Академии наук. 2015;461(2):1-3. [Sysoeva YuYu, Verbitskii EV. Kharakter aktivatsii mozga vo vremya nochnogo sna svyazan s lichnostnoi trevozhnost’yu individuuma. Doklady akademii nauk. 2015;461(2):1-3.(In Russ.)].
- Arapova YuYu, Shikhliarova AI, Verbitsky EV, Rostorguev EdE., Kuznetsova NS, Protasova TP. Electroencephalographic activity of wakefulness and sleep associated with primary brain tumors in human: a pilot study. J. Sleep Medicine and Disorders. 2016;:3(7):1068-1075.
- Вербицкий Е.В. Тревожность и сон. Журнал высшей нервной деятельности. 2013;63(1):6-12. [Verbitskii EV. Trevozhnost’ i son. Zhurnal vysshei nervnoi deyatel’nosti. 2013;63(1):6-12. (In Russ.)].
- Ковальзон В.М. Нейрофизиология и нейрохимия сна. Сомнология и медицина сна. Национальное руководство памяти А.М. Вейна и Я.И. Левина. Под ред. Полуэктова М.Г. М.: Медфорум; 2016: 11-55. [Koval’zon VM. Neirofiziologiya i neirokhimiya sna. Somnologiya i meditsina sna. Natsional’noe rukovodstvo pamyati A.M. Veina i Ya.I. Levina. Pod red. Poluektova M.G. M.: Medforum; 2016: 11-55. (In Russ.)].
- Середин С.Б., Воронина Т.А., Незнамов Г.Г., Жердев В.П. Феназепам: 25 лет в медицинской практике. М.: Наука; 2007. [Seredin SB, Voronina TA, Neznamov GG, Zherdev VP. Fenazepam: 25 let v meditsinskoi praktike. M.: Nauka; 2007. (In Russ.)].
- bingapis.com/images/search



