Регуляция лимфоцитами клеточного роста соматических тканей и новая иммунная теория старения
СтатьиОпубликовано в журнале:
Профилактика старения »» Выпуск 1 1998 (Обзор литературных и собственных данных)
В.И. Донцов
Национальный Геронтологический Центр
Теоретическое моделирование закономерностей процессов саморегуляции клеточного роста в организме указывает на необходимость существования специализированных клеточных популяций, выполняющих функцию межклеточной регуляции роста разнотипных соматических тканей. Анализ экспериментальных данных иммунологии и клеточной биологии показывает, что такая система может быть представлена специализированными Т-лимфоидными клетками. Снижение функции таких регуляторных клеток с возрастом может являться центральным механизмом для старения самообновляющихся соматических тканей организма и определять снижение с возрастом ростового потенциала тканей старого организма.
1. ПРОЦЕССЫ САМООРГАНИЗАЦИИ И САМООБНОВЛЕНИЯ ТКАНЕЙ ТРЕБУЮТ НАЛИЧИЯ СПЕЦИАЛИЗИРОВАННОЙ КЛЕТОЧНОЙ СИСТЕМЫ РЕГУЛЯЦИИ РОСТА ТКАНЕЙ
Нет сомнения, что проблема регуляции клеточного деления и тканевого роста является одной из главнейших как для теории, так и практики современой биологии и медицины. Современные теории старения также уделяют важнейшую роль в развитии процесса старения механизму снижения с возрастом клеточного самообновления тканей вследствие выраженного снижения их потенциала роста (1, 4, 5, 9, 11, 15, 16, 19, 22 - 24, 27).
Высоко организованные многоклеточные организмы состоят из огромного числа делящихся клеток, составляющих различные ткани и органы, которые растут и самообновляются различными темпами. Таким образом, само существование многоклеточных связано с надежными системами координации и взаимосогласования процессов клеточного деления и тканевого роста, знание которых на настоящий момент нельзя признать удовлетворительным.
Для рассмотрения процессов регуляции сложных разивающихся систем в настоящее время существует адекватный теоретический аппарат - это теории самоорганизации, разрабатываемые, например, в рамках синергетики и кибернетики. Прежде всего это работы нобелевских лауреатов: И. Пригожина, М.Эйгена и других ученых, работающих в этой области (14, 19, 25, 28, 29, 33, 40).
Одним из важнейших понятий в этой области в настоящее время является "гиперцикл" - он определяется как "принцип естественной самоорганизации, обусловливающий интеграцию и согласованную эволюцию системы функционально связанных самореплицирующихся единиц" (33).
Основой теории гиперцикла служит доказательство неизбежности формирования в процессе эволюции функциональных регуляторных связей более высокого порядка между самореплицирующимися единицами - системами более низкого порядка (например, между клетками или клеточными популяциями), что и формирует новую единую надсистему более высокого уровня. Моделирование таких взаимоотношений возможно с применением во многих случаях простых и наглядных схем, формул и графиков, при этом выявляются существенные моменты как общего плана, так и частного, если речь идет о рассмотрении конкретных физиологических процессов.
Рассмотрим некоторые общие представления такого рода.
Клеточный рост можно представить формально как движение клеток из фазы покоя (G) в фазу пролиферации (S) и затем в результате роста и деления надвое появление уже двух новых G клеток: G------S-----2G
В наиболее общем виде клеточное деление можно моделировать как рост числа клеток определенного типа (например, G), что, при отсутствии других влияний приводит только к бесконечному росту числа клеток по закону: dG/dt=k G, где "k" - коэффициент (т.е. изменение числа клеток пропорционально их числу в каждый момент времени).
Так как реально бесконечный неограниченно увеличивающийся процесс клеточного роста очень быстро приведет к гибели реального многоклеточного организма, должен существовать процесс ограничения числа клеток, зависящий от их числа, что в общем виде известно как обратная отрицательная связь: dG/dt = k1 G / (k2 G + C); ("k1", "k2" - коэффициенты, "c" - постоянная)
Именно такими рассуждениями были получены представления о КЕЙЛОННОЙ системе саморегуляции, работающей для одной клеточной популяции. При этом рост клеточной популяции оказывается ограниченным, а растущие клетки подавляют процесс входа в фазу роста покоящихся клеток: G -- (-)--> S
Дальнейшее рассмотрение, по существу, можно свести к аналогичным процессам самоорганизации, но уже для НЕСКОЛЬКИХ РАЗЛИЧНЫХ клеточных популяций. В этом случае каждая популяция клеток, имеющая свою кейлонную систему, рассматривается как отдельная элементарная система, и уже между ними будет идти процесс самоорганизации, который, принципиально, подчиняется ТЕМ же законам (формулам) - но под "G" теперь понимается уже определенная клеточная популяция в организме (например, гепатоциты или клетки эпидермиса, слизистых и т.п.).
В формуле число клеток dG будет пропорционально теперь количеству клеток популяции, влияющей стимулирующе и обратно пропорционально количеству клеток, влияющих ингибирующе на данную клеточную популяцию. Конкретный вид формул здесь может несколько различаться, но все они, как показавает анализ, дают принципиально одни и те же результаты (13 - 16), которые и представлены в общем виде далее. Все данные рассуждения следуют, впрочем, достаточно прямо из приведенных логических схем и для качественного понимания происходящих процессов не требуют расчетов и собственно математического моделирования.
Минимальное число клеточных популяций, составляющих КЛЕТОЧНЫЙ ГИПЕРЦИКЛ (назовем так такую надсистему), таким образом, три: две стимулируют друг друга (Ga и Gh) и одна ингибирует рост всех остальных (Gi):
Gi -- (+)--> Ga -- (+)--> Sh
-----------------------------
Si -- (+)--> Sb -- (+)--> Gh
dGa/dt = k1 Gh - k2 Gi - kа Ga
dGh/dt = k3 Ga - k4 Gi - kh Gh
dGi/dt = k5 (Sa+Sh) - ki Gi
(кейлонная система не показана, последний элемент формул учитывает спонтанную гибель клеток каждой популяции)
Такой клеточный гиперцикл имеет характерные черты - самостановление, саморегуляцию и самовосстановление (собственно, на данном этапе рассмотрения это одно и то же).
На Рис.1 и 2 показано компьютерное моделирование такого клеточного гиперцикла: самостановление из случайного начального состояния и восстановление после повреждения одного из элементов.
Видно, что все компоненты клеточного гиперцикла реагируют согласованно на внешние воздействия как единое целое.
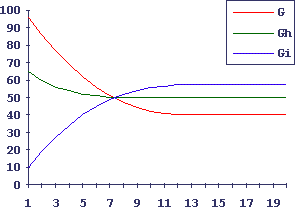
Рис.1. Самостановление клеточного гиперцикла, состоящего из 3-х клеточных популяций, из случайного начального положения
По оси абсцисс - число циклов клеточного деления; по оси ординат - число клеток в популяции.
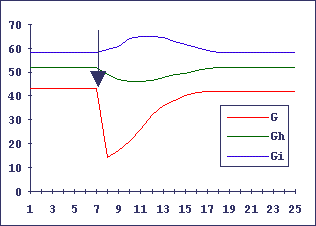
Рис.2. Самовосстановление (регенерация) клеточного гиперцикла при повреждении одного из компонентов
Обозначения как на предыдущем рисунке 1. Момент повреждения указан стрелкой.
Из общих законов биологии ясны дальнейшие направления эволюции такого КЛЕТОЧНОГО ГИПЕРЦИКЛА:
- увеличение числа клеточных популяций в составе гиперцикла ;
- специализация каждой популяции клеток:
б) в направлении регуляции клеточного роста - выделение регуляторных клеток стимулирующих (h) и ингибирующих (i) популяций в отдельную систему - назовем ее система клеточной регуляции пролиферации (КРП-система) - Gh, Gi и появление специфических клеток-регуляторов для клеток печени, почек и других регулируемых популяций,
в) выделение и усложнение регуляторных взамовлияний внутри самого клеточного гиперцикла - внутренние регуляторные связи.
На языке иммунологии с использованием теорий "регуляторных сетей" это можно представить как положительную селекцию специфических клонов, например, лимфоцитов, в ходе идиотипорганных взаимодействий, причем такие взаимоотношения могут быть распространены и на саму регуляторную систему с усложнением регуляторных взаимосвязей.
В конечном виде схема КРП-системы должна также учитывать появление в эволюции фазы глубокого покоя (Go) для клеток функциональных органов и тканей, так как выполнение функции приобретает для целостного организма важнейшее значение; в то же время для регуляторных клеток КРП-системы появление такой фазы только осложнило бы их функцию. В общем случае окончательно межклеточная регуляторная КРП-система может быть представлена в следующем виде (кейлонная система регуляции не показана):
ОБЩАЯ ПОЛНАЯ СХЕМА СИСТЕМЫ КРП
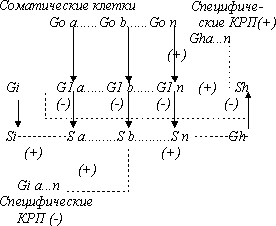
Математическое моделирование и схемы самоорганизации, самовосстановления и функционирования такой системы при различных процессах роста соматических клеток, как и пути становления этой системы в эволюции требуют специального рассмотрения и разобраны нами в отдельных публикациях (14 - 16, 27, 28), здесь же следует подчеркнуть прежде всего, что такая система в целом устойчива и обладает свойствами самостановления, самовосстановления и выхода в стационарное состояние при любом возмущении - внешнем воздействии и обладает, таким образом, свойством целостности и устойчивого самосохрания.
Т-ЛИМФОЦИТЫ - РЕГУЛЯТОРЫ РОСТА СОМАТИЧЕСКИХ КЛЕТОК
Основным вопросом является какие же именно клетки у современных млекопитающих и человека являются главными в регуляции роста соматических клеток организма. На этот вопрос есть совершенно четкие указания - как теоретические представления, так и экспериментальные данные указывают на преимущественную роль в таких процессах Т-лимфоцитов-регуляторов (хелперов и супрессоров), а именно, их ближайших предшественников, несущих SC-антиген (видимо, для неспецифических КРП) и Т-клеток, участвующих в так называемой "сингенной смешанной культуре лимфоцитов", которые реагируют "на свое", а не как иммунные клетки - на "чужое" (для специфических КРП).
Действительно, не трудно видеть при анализе требований, вытекающих теоретически из выше разобранных схем, что необходимые свойства, которыми должны обладать такие КРП, совпадают со свойствами регуляторных популяций лимфоцитов (главным образом Т-лимфоцитов).
КРП должны:
- быть подвижными (рециркулировать) в организме,
- иметь рецепторы к "своему" - осуществлять распознавание "своего",
- КРП должны обладать собственной пролиферацией, ускоряющейся в ходе выполнения ими функций регуляции клеточного роста других тканей,
- регуляция клетками КРП пролиферации должна иметь место для ЛЮБЫХ типов клеток в организме - в т.ч. лимфоцитов-эффекторов при иммунном ответе и пр.
Напомним главные экспериментальные факты из этой области:
наибольший международный и отечественный резонанс имели работы по переносу лимфоцитами "РЕГЕНЕРАЦИОННОЙ ИНФОРМАЦИИ":
- лимфоциты от животных с регенерацией тканей любого типа были способны индуцировать митозы и рост клеток соответствующего типа при сингенном переносе интактным животным (2, 12, 20, 26, 31, 32);
- перенос гиперпластической реакции лимфоцитами возможен для любых тканей и при любых процессах - например, при изопротеренол-индуцированной гипертрофии слюнных желез грызунов, при функциональной гиперплазии сердца и др. (8, 13, 20, 30, 31);
- известна регуляция Т-лимфоцитами пролиферации фибробластов в т.ч. выделяемых лимфокинов, специфических для них (1, 5, 6, 15);
- было показано, что симптомы разрастания костной ткани - остеопетроза, коррегируются переносом лимфоцитов от здоровых животных; более того, паратиреоидный гормон, специфический для остеокластов, действует через Т-лимфоциты, на которых, но не на остеокластах, имеются к нему рецепторы (36, 39);
- было показано, что общую задержку роста - карликовость мышей можно ликвидировать переносом лимфоцитов от здоровых животных, а на Т-лимфоцитах есть рецепторы к соматотропному гормону и число их выше в период роста животных (21, 24);
- известен парадокс "Nude" мышей, противоречащий теории опухолевого надзора, (опухолевый надзор видвигался как главный эволюционный фактор становления лимфоидной системы): у бестимусных мышей частота спонтанных и индуцированных опухолей не повышена, а снижена; более того, повышение частоты опухолей у них восстанавливается при переносе лимфоцитов от обычных мышей; у этих же животных снижение регенерации и самообновления тканей (дистрофический синдром) не может быть объяснен только снижением иммунитета и бактериальной инфекцией; у этих же мышей со временем Т-лимфоциты все равно появляются, то есть, роль тимуса играют обычные ткани, также стимулирующие пролиферацию и созревание Т-клеток (21);
- известны факты стимулирования и ингибирования лимфоцитами роста опухолей, не сводящиеся к типичным чисто иммунным феноменам (1, 4, 10, 15, 21, 22, 34, 38);
- при старении снижению клеточного самообновления (физиологической регенерации) предшествует дегенерация тимуса - источника Т-лимфоцитов (4, 5, 15, 21, 23);
- реакция "трансплантат против хозяина" и иные методы, приводящие к инактивации собственных лимфоцитов "хозяина" сопровождаются типичными "неиммунными" симптомами - снижением индуцированной регенерации, дистрофией тканей и пр. (5, 15, 21);
- система Т-клеток-регуляторов гораздо более сложна, чем система Т- и В-эффекторов иммунитета, причем Т-клетки иммунной системы распознают чужеродный антиген ("чужое") в комплексе со "своим" - антигенами главного комплекса гистосовместимости (5, 15, 34);
- подробно описана и исследована в иммунологии т.н. "сингенная смешанная культура лимфоцитов" - sMLC, когда Т-клетки преимущественно Т-хелперного и Т-супрессорного (но не киллерного) типов реагируют бурной пролиферацией на СВОИ клетки организма, в том числе на неиммунные клетки; в классической иммунологии эти клетки - "лишние" - их можно убрать антисыворотками и обычные иммунные реакции не изменятся ; в рамках sMLC интересны факты появления или повышения количества Ia рецепторов, на которые реагируют
- Т-клетки, на неиммунных клетках различных тканей при подготовке их к клеточному делению, в том числе на опухолевых клетках и пр. (15, 34, 35, 37, 38).
Известно из иммунологии и иммуногенетики рецепторов Т-клеток, что "распознавание чужого" в принципе не отличается от "распознавания своего", причем возможен перевод "своего" в "чужое" (аутоиммунные реакции) и "чужого в свое" (толерантность) в результате прежде всего изменений в регуляторных Т-клетках.
При распознавании Т-зависимых молекулярных антигенов в ходе иммунной реакции "чужое" распознается в комплексе со "своим" - собственными антигенами главного комплекса гистосовместимости, которые представлены на "презентирующих"антиген клетках разного типа. Необходимость "презентации" антигена для лимфоцитов Ia-несущими клетками собственного организма объясняется тем, что эволюционно Т-клетки приспособлены к реакции именно на клетки, причем на готовящиеся к пролиферации клетки, а не на молекулы.
Исходя из всех выше приведенных фактов, мы предположили, что функция регуляции клеточного роста "своих" клеток является филогенетически более древней и более важной - собственно это и есть эволюционная сила, формирующая сложную систему Т-лимфоцитов-регуляторов пролиферации любых клеток, в том числе Т- и В-эффекторов иммунитета, которые филогенетически более поздние и более простые. В таком случае, иммунная система - только специализировавшаяся в отношении элиминации "чужого" часть более сложной и общей системы - системы регуляции клеточного роста - КРП системы (14, 15).
Можно сделать некоторые очевидные выводы:
1. Должна существовать на межклеточном уровне специальная система регуляции роста соматических тканей, появившаяся, сложившаяся и усложняющаяся очень рано в эволюции.
2. Элементами такой системы являются КРП - некоторые популяции Т-лимфоцитов (для современных млекопитающих).
3. Элементарные свойства клеток такой системы можно видеть и для иных типов клеток, что позволяет проследить становление такой системы в эволюции (это прежде всего макрофаги и большие гранулярные лимфоциты).
4. Через данную систему осуществляются многие регуляторные влияния иерархически более высокого системного уровня (например, гормонального и еще более иерархически высокого - нервного).
5. Функционирование такой системы четко описывается и моделируется.
6. Именно необходимость регуляции клеточного роста в сложном организме, а не надзор за опухолями и явилась эволюционной причиной формирования сложной Т-лимфоидной системы, которая в иммунологии рассматривается односторонне как регуляторная система только для иммуноцитов.
7. КРП можно выделять, изучать, влиять на их активность, получать регуляторные специфические и неспецифические факторы и через них влиять на различные физиологические и патологические процессы.
8. КРП-система является особой, ОТДЕЛЬНОЙ от иммунной, системой организма, решающей задачи более общего и важного плана, чем собственно иммунная. Иммунная система может быть сведена к специализировавшейся части КРП-системы.
Осознанное отношение к лимфоцитам-регуляторам роста соматических клеток как к элементам КРП-системы позволило нам получить качественно более значимые результаты как при теоретическом рассмотрении, так и при экспериментальной работе.
В частности, нами были выделены и охарактеризованы такие Т-регуляторы роста различных соматических клеток организма, изучена их кинетика, особенности фенотипа, реакция на некоторые фармакологические агенты, выделение регуляторных факторов, специфических для соматических клеток и др. (8, 10, 11, 13-16).
Для планирования экспериментального изучения системы КРП чрезвычайно полезным оказывается теоретическое рассмотрение вероятных закономерностей функционирования системы КРП.
Индукция КРП-системы - выход ее из динамического равновесия, исходя из общей схемы, возможна по следующим типам:
1. Регенерационный запуск (Go - G1 механизм, запускаемый естественными регуляторами той или иной ткани при ее повреждении),
2. Перенос регуляторных клеток (Gh и Gi - механизм) - искусственный механизм, использованный, например, А.Г. Бабаевой при воспроизведении феномена "переноса регенерационной информации" (1, 2),
3. Функционально-гипертрофический запуск (Go - G1 механизм), например, воспроизводимый Г.Я. Свет-Молдавским с соавторами при изучении пассивного переноса лимфоидными клетками компенсаторной гипертрофии миокарда (30),
4. Экспериментальный фармакологический запуск (Go - G1 механизм), воспроизведенный, например, нами при изопротеренол-индуцированной гипертрофии слюнных желез грызунов (5, 8, 13, 14),
5. Экспериментальный фармакологический запуск (Gh и Gi - механизм по типу пункта 2), также полученный нами на аналогичной модели (2, 5, 13, 14),
6. Интерференция при любом клеточном росте (регенерация, опухолевый рост, иммунная реакция) за счет неспецифических КРП (Gh и Gi - механизм) - неоднократно показанные феномены интерференции процессов роста различных типов клеток, в том числе иммунокомпетентных клеток.
Все эти типы индукции системы КРП сводятся, как нетрудно видеть, всего к двум типам запуска:
а) повышение числа клеток в G1 состоянии;
б) изменение активности и количества самих КРП клеток.
Исходя из известных свойств Т-лимфоцитов, которым приписывается роль КРП, а также исходя из выше приведенных теоретических схем, можно также ожидать и более частных особенностей функционирования системы КРП, которые действительно наблюдаются во многих типах экспериментов самых различных авторов:
- быстрая (в течение часов и минут) активация КРП/+/,
- это возможно если у КРП/+/ нет Go фазы - они должны быть заторможены в G1/S стадии (филогенетически древний механизм),
- при любом росте соматических клеток идет пролиферация и КРП/+/,
- выделение клетками КРП/+/ регуляторных стимулирующих клеточный рост факторов;
- реакция на "свое" (Н-2 зависимость эффекта) для неспецифических КРП (филогенетически древний механизм);
- в иммунологии должна быть сингенная смешанная культура лимфоцитов "на свое" (действительно есть sMLC);
- поздняя активация КРП/-/ - уже после начала пролиферации соматических клеток;
- КРП/-/ должны пролиферировать быстрее чем КРП/+/, видимо, постоянно медленно пролиферировать и увеличивать скорость пролиферации в ходе своей активации;
- КРП/-/ должны выделять регуляторные ингибирующие рост факторы,
- КРП могут мигрировать в организме в ходе активации и функционирования;
- при вторичном ответе должны преобладать специфические КРП/+/ и КРП/-/,
- специфические КРП должны иметь случайный клоновый механизм активации (реагировать не "на свое", а "на все" в том числе "на чужое" - они могут быть - Н-2 не зависимы);
- КРП специфические могут случайно формировать клоны к "чужому", что дает возможность развития иммунитета как специализации системы КРП к антигену,
- в иммунной реакции сохраняются закономерности КРП-реакция "на свое", взаимодействие специфических и неспецифических КРП и другие характерные для системы КРП свойства,
- между различными процессами с клеточной пролиферацией должна быть интерференция (регенерация - опухолевый рост - иммунитет и другие процессы интерференции),
- при возрастном снижении активности КРП снижается и уровень фонового потенциала роста тканей, но это регуляторный и принципиально обратимый процесс,
- при опухолевом росте активируются КРП/-/ и в меньшей степени КРП/+/,
- так как регуляция опухолевых клеток нарушена, то при опухолевом росте подавляется регенерация, иммунитет и пр., за счет избыточной активации КРП/-/ - системы,
- КРП имеет СВОИ СПЕЦИФИЧЕСКИЕ задачи и формирует СВОЮ СПЕЦИАЛЬНУЮ СИСТЕМУ в организме, отличную от иммунной и филогенетически более древнюю, решающую более важные задачи, чем иммунная система - задачи ИНТЕГРАЦИИ многоклеточного сообщества в единый организм,
- иммунитет появляется как специализация ЧА-СТИ системы КРП в иммунную в ходе реакции "на чужое" (в филогенезе и онтогенезе).
Рассмотрим только некоторые экспериментальные примеры, полученные нами на крысах Wistar, August и мышах BALB/c, C57Вlack/6 и CВА, цель которых - показать возможность и диапазон применения разрабатываемых теоретических и практических подходов. Детали экспериментов представлены в подписях под рисунками.
На Рис.3 показана кинетика формирования бластных клеток селезенки (в процентах к контролю у интактных животных) в ходе индукции регенерации печени после повреждения гепатоцитов четыреххлористым углеродом - типичная модель, на которой был подтвержден феномен переноса регенерационной информации (32). Внизу для сравнения даны периоды клеточного цикла для гепатоцитов.
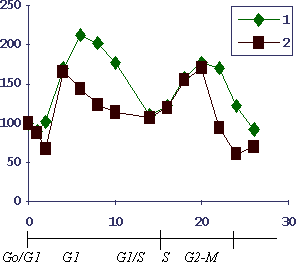
Рис.3. Кинетика формирования бластных клеток селезенки при индукции четыреххлористым углеродом регенерации печени у мышей
По оси абсцисс - время в часах после введения перорально 0,2 мл. 0,15% масляного раствора ССl4; по оси ординат - параметры в процентах к интактным животным. Здесь и далее представлены обычно средние значения по 5-6 животным. 1 - количество бластных клеток селезенки, не осаждающихся при центрифугировании в градиенте плотности фиколла 1.07. 2- включение 3Н-тимидиновой метки в кислото-нерастворимую фракцию (ДНК) 4-х часовой культуры переживающих клеток селезенки.
Видны типичные для процессов такого рода теоретически предсказанные 2 пика формирования бластных клеток Т-лимфоидной природы, как было показано в тестах с анти-Тhy1 и анти-тимоцитарной сывороткой и комплементом.
Перенос клеток таких пиков in vivo интактным мышам вызывает стимулирование (первый пик) или снижение (второй пик) митозов печени и изменяет включение 3Н-тимидина в интактной или регенерирующей печени реципиентов (15).
Аналогичная кинетика наблюдается при феномене индуцированной бэта-адреномиметиками гиперплазии слюнных желез у грызунов - феномене Селье. Это хорошо изученный и достаточно широко известный феномен увеличения массы слюнных желез грызунов в результате массовой индукции фармакологическими средствами Go/G1 перехода клеток этих желез с последующим синтезом ДНК, делением и ростом клеток желез (3, 15): Рис. 4 и 5.
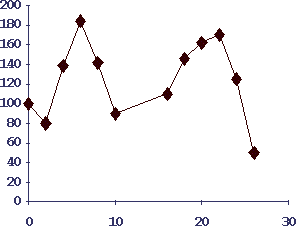
Рис.4. Кинетика формирования бластных клеток селезенки при изопротеренол-индуцированной гиперплазии подчелюстных слюнных желез у мышей
По оси абсцисс - время в часах после внутрибрюшинного введения мышам 5 мг изопротеренола; по оси ординат - количество бластных клеток селезенки (плотностью мнее 1.07). Видны те же два пика активации лимфоцитов как в селезенке, так и в лимфоузлах шеи.
Сингенный перенос активированных клеток I пика интактным реципиентам повышает у них массу слюнных желез (8, 15).
Можно разделить собственно индукцию Go/G1 перехода адреномиметиками и G1/S индукцию перехода лимфоцитами, вводя циклофосфан или гидрокортизон реципиентам, т.е., создавая у них иммунодефицит, что ведет к отмене гиперпластической реакции слюнных желез у иммунодефицитных мышей, тогда как переносимые лимфоциты восстанавливают эту гиперплазию (3, 15).
С использованием антисывороток против Т-лимфоцитов можно убедиться, что с течением времени среди активированных бластных лимфоидных клеток увеличивается количество зрелых, несущих Thy1 антиген, типичный для Т-лимфоцитов, и снижается число несущих SC-антиген, типичный для предшественников Т-лимфоцитов (15).
Такие антисыворотки отменяют процесс гиперплазии ткани как при введении in vivo, так и при обработке лимфоцитов в ходе сингенного переноса интактным животным.
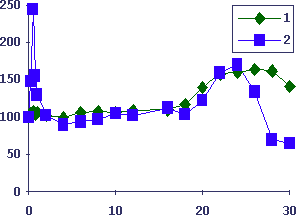
Рис.5. Кинетика формирования бластных клеток подчелюстных лимфоузлов при изопротеренол-индуцированной гиперплазии подчелюстных слюнных желез у мышей
Обозначения как на Рис. 4. 1 - масса подчелюстных слюнных желез, 2 - количество бластных клеток лимфоузлов (плотностью менее 1.07)
Таким образом, неспецифические КРП, активированные в ходе первичной реакции стимуляции роста соматических тканей, представлены предшественниками Т-лимфоцитов, несущими SC-антиген, которые стимулируются к пролиферации и созреванию сами и активируют рост соматических тканей (в нашем случае - ткани печени и слюнных желез).
При переносе активированных бластных клеток селезенки и лимфоузлов интактным или изопротеренол-обработанным мышам были показаны характерные для КРП особенности (Рис. 6 и 7):
- наличие клеток памяти КРП (отличие КРП, реагирующих при первом и последующих воспроизведениях реакции гиперплазии ткани),
- сингенное ограничение для первичной реакции и Н-2 не ограниченность эффекта для КРП памяти,
- наличие неспецифических КРП, ограниченных по Н-2 системе гистосовместимости и ряд других особенностей, показанных ниже.
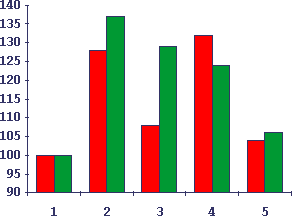
Рис.6. Отличия специфических и неспецифических КРП/+/ в тесте сингенного и аллогенного переноса активированных лимфоцитов у мышей
По вертикали - интенсивность гиперплазии слюнных желез у мышей через сутки после внутривенного переноса 10 млн. активированных бластных клеток I пика от мышей-доноров КРП, получивших изопротеренол или ССl4; по горизонтали - условия переноса. Темные столбики - однократное введение веществ; светлые - двухкратное введение веществ. 1 - контроль интактных животных (принят за 100%), 2 - перенос клеток от сингенных доноров, получивших изопротеренол, 3 - то же в аллогенной системе, 4 - перенос клеток от сингенных доноров, получивших ССl4, 5 - то же в аллогенной системе.
Рост ее сопровождается увеличением массы селезенки, числа бластных клеток в селезенке и повышением включения 3Н-тимидина в лимфоциты селезенки - Рис. 8.
У иммунодефицитных животных опухоль не растет, но рост ее может быть восстановлен введением сингенных Т-клеток от мышей-опухоленосителей или от мышей с индукцией роста иных типов тканей (10, 15).
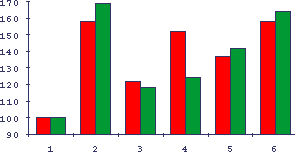
Рис.7. Отличия специфических и неспецифических КРП/-/ в тесте сингенного и аллогенного переноса активированных лимфоцитов у мышей
Обозначения как на Рис.6. Перенос производился для активированных бластных клеток КРП - II пика.
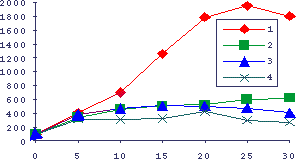
Рис.8. Активация лимфоидной ткани при росте опухоли АКАТОЛ у мышей
По оси абсцисс - время в днях после подкожной перевивки 30 мг ткани АКАТОЛ, по оси ординат - параметры в процентах к интактным животным. 1 - масса опухоли, 2 - масса селезенки, 3 - бластные клетки селезенки, 4 - включение 3Н-тимидина в ДНК клеток селезенки.
Исходя из общей теоретической схемы должны существовать стимулирующие и ингибирующие КРП неспецифического типа - видимо, самые древние типы КРП. Их можно легко обнаружить в процессах интерференции при индукции роста двух различных тканей, например, при воспроизведении реакции гиперплазии слюнных желез у мышей с регенерацией печени (Рис.9) или при воспроизведении той же реакции у мышей-опухоленосителей. В свою очередь, рост опухоли у мышей мог быть подавлен при введении одновременно с перевивкой опухоли клеток II пика (неспецифические КРП/-/) от мышей, получивших инъекцию изопротеренола, ССl4 или от мышей с развившейся уже опухолью (Рис.10).
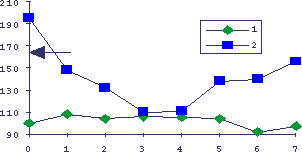
Рис.9. Интерференция процессов регенерации печени и гиперплазии слюнных желез у мышей
По оси абсцисс - время в днях после индукции регенерации печени введением ССl4, по оси ординат - параметры в процентах к интактным животным. 1 - масса слюнных желез у мышей, не получавших изопротеренол, 2 - гиперпластическая реакция на изопротеренол у мышей с регенерацией печени. Стрелкой показана реакция на изопротеренол у интактных животных.
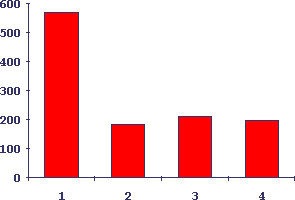
Рис.10. Подавление роста опухоли у мышей введением КРП/-/ от животных с индукцией тканевого роста разного типа
По оси абсцисс - условия переноса, по оси ординат - рост опухоли в процентах от перенесенной опухолевой массы на 7-й день после перевивки опухоли. 1 - контроль опухолевого роста, 2 - перенос бластных клеток от мышей-опухоленосителей, 3 - перенос бластных клеток селезенки от мышей с индукцией регенерации печени, 4 - перенос бластных клеток селезенки от мышей с индукцией гиперплазии слюнных желез.
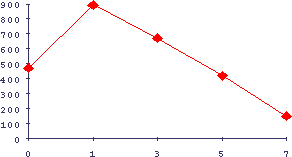
Рис.11. Интерференция процессов иммунной реакции и регенерации печени
По оси абсцисс - время после индукции регенерации печени введением ССl4, по оси ординат - количество иммунных бляшек на 1 млн клеток селезенки на 4-й день иммунизации эритроцитами барана в тесте по Каннингаму.
Возможно подавление в сингенном переносе роста опухоли АКАТОЛ при переносе мышам-опухоленосителям лимфоцитов от:
а) мышей-опухоленосителей;
б) от изопротеренол-обработанных или
в) ССI4-обработанных мышей (лимфоциты необходимо брать в точно определенные сроки - второй пик активированных КРП).
Выявляется по общему механизму, разобранному выше, также четкая интерференция процессов регенерации печени и иммуной реакции на эритроциты барана (Рис.11).
Специфичность лимфоцитов к определенной ткани можно показать в обычном розеточном тесте с эритроцитами, нагруженными мембранами таких тканей.
Таблица 1. СВОДНАЯ ТАБЛИЦА ФЕНОТИПИЧЕСКИХ СВОЙСТВ КРП /+/ и КРП/-/
| N | СВОЙСТВА | КРП/+/неспец. | КРП/-/неспец. | КРП/+/специфич. | КРП/-/специфич. |
| 1. | Сроки активации | 2-6 часов | 20-30 часов | II-реакция | II-реакция |
| 2. | Состояние в покое | в G1 состоянии | в S состоянии | в Go ? | в Gо ? |
| 3. | Характер активации | запуск G1/S | актив.пролиф. | клоновый рост | клоновый рост |
| 4. | Чувствительность | к G1 кл-мишени | к S состоянию | к АГ тканей | к АГ тканей |
| 5. | Антигены | SC-Thy1, Ly1 | Тhy1, Ly2 | больше Ly2 | больше Ly2 |
| 6. | Н-2 специфичность | есть | есть | не типична | не типична |
| 7. | Ткане-специфичны | нет | нет | да | да |
| 8. | Предшественники | замена 3-4 дня | замена 24 ч. | длительно | длительно |
| 9. | Интерференция | неспец.стимул. | неспец.ингиб. | cпециф.память | специф.память |
| 10. | Иммунная память | не типична | не типична | типична | типична |
Таблица 2. СВОДНАЯ ТАБЛИЦА МОЛЕКУЛЯРНО-БИОЛОГИЧЕСКИХ СВОЙСТВ КРП/+/ и КРП/-/
| N | СВОЙСТВА | КРП/+/неспецифические | КРП/-/неспецифические |
| 1. | Плотность активированных клеток | 1.07 | 1.06 |
| 2. | Актиномицин Д (синтез РНК) | подавляет | мало эффективен |
| 3. | Циклофосфан (синтез ДНК) | умеренная чувствительность | высокая чувствительность |
| 4. | Ингибиторы протеаз | подавляет | мало эффективен |
| 5. | Цинк хлорид (Са-зависимость) | подавляет | подавляет |
| 6. | Гидрокортизон-чувствительность | умеренная | высокая |
| 7. | Простагландины: | ||
| ПГЕ2 | |||
| рецепторы | есть | есть | |
| эффект на пролиферацию | стимуляция | не влияет (?) | |
| синтез факторов регуляции | не влияет (?) | стимуляция | |
| ПГФ2-альфа: | |||
| рецепторы | есть | есть | |
| эффект на пролиферацию | подавление | не влияет (?) | |
| синтез факторов регуляции | стимуляция(?) | не влияет (?) | |
| 8. | Рецепторы к БАВ: | ||
| серотонин | есть | есть | |
| гистамин | есть (Н-1) | есть (Н-2) | |
| 9. | Ингибитора аминооксидаз: | ||
| иМАО | э ф ф е к т и в е н in vivo | ||
| иДАО | э ф ф е к т и в е н in vivo | ||
| 10. | ДОФА | э ф ф е к т и в е н in vivo | |
| 11. | Лимфокейлон | э ф ф е к т и в е н in vivo | |
| 12. | Литий хлорид | э ф ф е к т и в е н in vivo | |
| 13. | Эффект цАМФ: | ||
| на пролиферацию | стимуляция | не эффективен (?) | |
| рецепторы | перенос в клетку | --- | |
| Зависимость от Са: | |||
| внеклеточного | не зависит | --- | |
| внутриклеточного | зависит | --- | |
| зависимость от синтеза белка | зависит | --- | |
| 14. | Эффект цГМФ: | ||
| на пролиферацию | не эффективен(?) | стимуляция | |
| рецепторы | --- | на мембране | |
| Зависимость от Са: | |||
| внеклеточного | --- | зависит | |
| внутриклеточного | --- | не зависит | |
| зависимость от синтеза белка | --- | не зависит | |
Специфические КРП более четко проявляли свои эффекты при повторных индукциях роста соматических тканей, при этом обнаруживался эффект "памяти" и отличия специфических КРП от неспецифических, например, отсутствие Н-2 ограничения их эффекта (2, 15).
Факторы, выделяемые КРП, можно очистить стандартными методами гель-хроматографии и изоэлектрофокусирования.
Адсорбцией на активированной сефарозе с привязанными выделенными мембранами тканей можно показать наличие специфических и неспецифических факторов подобного типа (15)
Наконец, КРП - бластные клетки пиков, можно исследовать средствами клеточной и молекулярной иммунологии и биологии: изучить их фенотип, дифференцировку, рецепторы, механизмы активации, чувствительность к различным веществам и пр., что и было во многих отношениях сделано (1, 2, 6, 10, 13, 14, 16, 20, 22, 23, 24, 26, 27, 28, 30,32, 34, 35, 37, 38, 39).
Здесь приведены таблицы первичной оценки свойств КРП /-/ и/+/ специфического и неспецифического типа, полученные нами в ходе ряда экспериментов (8-11, 14, 15, 16, 19, 27, 28).
Учитывая значимость снижения потенциала клеточного роста соматических тканей для процессов старения многоклеточных организмов, нами была сформулирована новая иммунная теория старения, указывающая на важную роль в развитии процессов старения возрастного дефицита лимфоцит-зависимой функции регуляции роста соматических тканей.
ВОЗРАСТНОЙ ДЕФИЦИТ ЛИМФОЦИТ-ЗАВИСИМОЙ РЕГУЛЯЦИИ РОСТА СОМАТИЧЕСКИХ ТКАНЕЙ КАК ЦЕНТРАЛЬНЫЙ МЕХАНИЗМ СТАРЕНИЯ МНОГОКЛЕТОЧНЫХ
После формулировки Ф.Бернетом иммуноло-гической теории старения функции иммунной системы в период старения подверглись пристальному изучению, была показана несомненная связь функций иммунной системы с процессом старения (5, 13, 16, 19, 21, 23, 27, 28):
- прогрессирующая атрофия тимуса и всей лимфоидной ткани с возрастом,
- снижение предстоящей продолжительности жизни при снижении количества циркулирующих Т-лимфоцитов,
- сходство старческих изменений и процессов, наблюдаемых при ранней тимэктомии и иммунодефицитах иной природы, иммунологические расстройства и нарушения экспрессии антигенов гистосовместимости при прогериях и пр.
- атрофия тимуса, селезенки и лимфоузлов, снижение числа периферических Т-клеток,
- увеличение числа незрелых лимфоцитов вследствие задержки их дифференцировки,
- снижение числа предшественников Т-клеток и выраженное снижение продукции тимических гормонов с диссоциацией в сторону активации Т-супрессорных механизмов, сопровождающейся активацией неспецифических Т-хелперов и Т-супрессоров, что растормаживает аутоиммунные процессы,
- уменьшение разнообразия антигенного репертуара лимфоцитов,
- снижение продукции ИЛ-2 Т-хелперами,
- выраженное снижение способности лимфоцитов активироваться собственными клетками организма - снижение так называемой сингенной смешанной культуры лимфоцитов,
- снижение противоопухолевой устойчивости организма и др.
Известно, что такое снижение самообновления разнообразных тканей с возрастом сопровождаeтся по неясным причинам развитием генерализованного G1/S блока для процессов пролиферации: увеличивается число готовых к пролиферации, но не получивших дальнейшего стимула для этого процесса клеток.
В то же время, в иммунологии хорошо известен такой G1/S блок, типичный для лимфоцитов-эффекторов старых животных, но в иммунологии хорошо изучена многими авторами и причина его - дисбалланс функций Т-лимфоцитов-регуляторов, заключающийся в снижении общего числа Т-регуляторов и повышении доли Т-ингибиторов.
Эти данные, наряду с известным резким снижением sMLC в старости, корреспондируют с выше разобранными представлениями о системе КРП в организме.
Исходя из всего этого, нами была разработана новая лимфоидная теория старения.
Основные ее положения могут быть сведены к следующему:
1. Ведущим механизмом старения соматических тканей является снижение их клеточного самообновления.
2. Снижение потенциала клеточного роста соматических тканей при старении определяется изменениями в системе лимфоидной регуляции пролиферации соматических клеток.
3. Сущностью изменений КРП в старости является увеличение доли КРП ингибиторов и абсолютное снижение общего числа КРП.
4. Нарушение соотношения КРП стимулирующего и ингибирующего типов ведет к снижению скорости продвижения соматических клеток из фазы G1 в S, формируя G1/S блок в тканях старых животных.
5. Прогрессирующее снижение пролиферативной активности клеток соматических тканей ведет к увеличению доли "старых" клеток, при этом "старческие" изменения являются результатом проявления нормальных свойств таких, углубившихся в состояние дифференцировки, клеток с длительным периодом жизни.
6. Так как в тканях всегда идут процессы замещения молодыми клетками, наряду с процессами старения формируются процессы приспособления, гипертрофии и пр.
7. Изменения системы КРП являются результатом продолжения действия регуляторов ограничения роста организма после того, как ост закончен, при участии гипоталамо-гипофизарной системы и тимуса.
Некоторые экспериментальные данные, подтверждающие эти выводы, представлены ниже.
Возрастной иммунодефицит показан на Рис.12. Видно, что с возрастом резчайшим образом снижается относительное количество лимфоидной ткани в отношении на единицу массы животного. Такой иммунодефицит сопровождается также выраженным снижением с возрастом потенциала роста тканей и снижением реакции КРП в ходе рост-индуцируемых процессов - Рис.13. При этом кинетика гиперпластической реакции изменяется характерным для старения образом: удлиняется время реакции, время достижения пика гиперпла-зии, снижается абсолютное значение достигаемого максимума реакции - Рис.14.
Этот иммунодефицит можно коррегировать некоторыми иммуномодуляторами - Рис.15. Эти же вещества способны восстанавливать и ростовые потенции тканей старых мышей - Рис.16. Известно, что соли лития и цинка обладают выраженным иммунокоррегирующим эффектом, влияя на обмен одно- и двувалентных ионов, на индукцию Т-клеток в ходе иммунной реакции. Типичен дозозависимый эффект этих веществ и благоприятное действие в пожилом возрасте, что определяет включение их в состав ряда биостимулирующих и иммунокоррегирующих фармакопрепаратов (15, 16 27, 28).
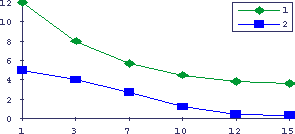
Рис. 12. Снижение относительной массы лимфоидных органов у старых животных
По оси абсцисс - возраст мышей BALB/c в месяцах, по оси ординат - относительная масса лимфоидных органов (мг массы органа на 1 г массы тела). 1 - относительная масса селезенки, 2 - относительная масса тимуса (х10).
Что касается ДОПА, то известно, что он является предшественником дофамина и вмешивается в обмен нейромедиаторов мозга, ответственных за иммунорегуляцию. Показано, в частности, что дофамин- и серотонин- эргические механизмы реципрокно регулируют иммунитет у млекопитающих (7) и могут включаться в механизмы индукции с возрастом процессов атрофии тимуса. В наших экспериментах ДОПА восстанавливал кинетику реакции на изопротеренол у старых мышей - Рис. 17.
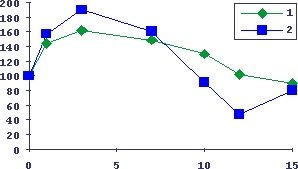
Рис.13. Возраст-зависимое снижение изопротеренол-индуцированной гиперплазии слюнных желез у мышей
По оси абсцисс - возраст мышей BALB/c в месяцах, по оси ординат - параметры в процентах к интактным животным. 1 - гиперпластическая реакция ткани слюнных желез, 2 - интенсивность бластообразования в селезенке.
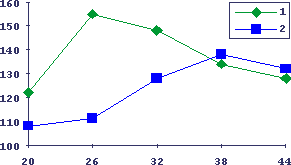
Рис.14. Изменение кинетики изопротеренол-индуцированной гиперплазии слюнных желех у мышей с возрастом
По оси абсцисс - время в часах после инъекции изопротеренола, по оси ординат - гиперпластическая реакция ткани слюнных желез, 1 - молодые мыши, 2 - старые мыши.
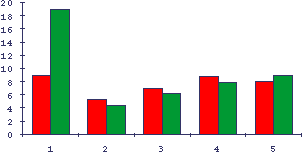
Рис.15. Влияние солей лития, цинка и ДОПА на массу лимфоидных органов у старых мышей
По оси абсцисс - условия, по оси ординат - относительная масса лимфоидных органов. Темные столбики - селезенка, светлые - тимус (х10). 1 - контроль молодых животных (возраст 3 месяца), 2 - контроль старых животных (возраст 12 месяцев), 3 - введение соли лития, 4 - введение соли цинка, 5 - введение L-DOPА.
Особый интерес представляет возможность восстановления ростового потенциала тканей старых мышей сывороткой крови от молодых животных, что отражено ниже - Рис. 18.
Наконец, мы показали, что сыворотка крови таких молодых животных была способна стимулировать пролиферацию клеток системы КРП - Рис. 19. Переживающие культуры клеток КРП усиливали включение 3Н-тимидина в присутствии в среде сыворотки крови молодых мышей.
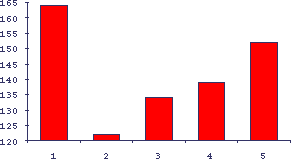
Рис.16. Влияние солей лития, цинка и ДОПА на изопротеренол-индуцированную гиперплазию слюнных желез у старых мышей
По оси абсцисс - условия, по оси ординат - реакция на изопротеренол в процентах к интактным животным. 1 - контроль молодых животных (воз-раст 3 месяца), 2 - контроль старых животных (возраст 12 месяцев), 3 - соль лития, 4 - соль цинка, 5 - L-DOPА.
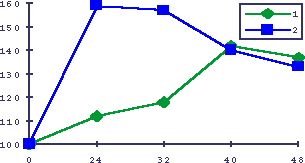
Рис. 17. Восстановление ДОПА кинетики изопротеренол-индуцированной гиперплазии слюнных желез у старых мышей
По оси абсцисс - время в часах после введения изопротеренола, по оси ординат - гиперпластическая реакция слюнных желез на изопротеренол в процентах к интактным животным. 1 - старые животные, 2 - то же после введения L-DOPА.
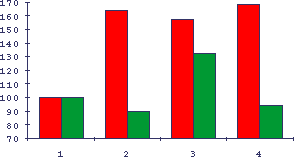
Рис.18. Восстановление сывороткой крови молодых мышей изопротеренол-индуцированной гиперплазии слюнных желез у старых мышей
По оси абцисс - условия, по оси ординат - гиперпластическая реакция слюнных желез на изопротеренол в процентах к интактным животным.Темные столбики - молодые животные, светлые - старые. 1 - интактные мыши (принято за 100%), 2 - введение изопротеренола, 3 - то же после введения внутривенно 0,3 мл сыворотки крови молодых мышей, 4 - то же для сыворотки крови старых мышей.
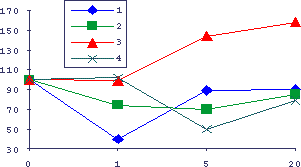
Рис.19. Влияние сыворотки молодых и старых мышей на пролиферацию клеток системы КРП
По оси абсцисс - концентрация сыворотки крови в переживающих 4-х часовых культурах бластных клеток селезенки, по оси ординат - включение 3Н-тимидина в кислотонерастворимую фракцию культур в процентах к культуре без добавления сыворотки. 1 - Молодые мыши и молодая сыворотка, 2 - молодые мыши и старая сыворотка, 3 - старые мыши и молодая сыворотка, 4- старые мыши и старая сыворотка.
Можно, таким образом, полагать, что при старении резко снижается функция Т-лимфоцитов-КРП системы как результат изменений в регуляторных системах организма. Мы обнаружили ряд таких данных экспериментально и показали возможность реактивации и быстрого восстановления потенциала роста клеток при воздействии на клетки КРП-системы (13 - 16).
Предлагаемая новая иммунная теория, таким образом, имеет не только теоретический интерес, но и позволяет использовать весь потенциал иммунофармакологии для противодействия одному из важнейших механизмов старения - снижению с возрастом клеточного самообновления у многоклеточных, в том числе у млекопитающих и человека.
ВЫВОДЫ
1. Существование многоклеточных организмов требует надежных регуляторных систем координации и взаимосогласования процессов клеточного деления и тканевого роста.
2. Важнейшим уровнем такой регуляции является система межклеточной регуляции клеточного роста разнотипных соматических тканей.
3. Теоретическое моделирование закономерностей самостановления и саморегуляции такой системы указывает на необходимость существования специализированных клеточных популяций, выполняющих функцию межклеточной регуляции роста соматических тканей.
4. Рассмотрение экспериментальных данных иммунологии и клеточной биологии показывает, что такая система может быть представлена специализированными Т-клетками - стимулирующих и ингибирующих популяций, реагирующих в "сингенной смешанной культуре лимфоцитов" на собственные растущие клетки организма.
5. Снижение функции таких Т-регулирующих клеток с возрастом может являться центральным механизмом для старения самообновляющихся соматических тканей организма и определять снижение с возрастом ростового потенциала тканей старого организма.
ЛИТЕРАТУРА
1. Бабаева А.Г. Традиционные и нетрадиционные представления о роли системы иммуногенеза в организме // Вестник АМН СССР. 1986. N 1. С.22.
2. Бабаева А.Г., Нестеренко В.Г., Юдина Н.В. Усиление способности лимфоцитов мышей с повторными резекциями печени к переносу регенерационной информации // Бюл. эксперим.
биол.мед. 1982. N 6. С. 98-99.
3. Бабаева А.Г., Шубникова Е.А. Структура, функции и адаптивный рост слюнных желез. М.:МГУ. 1979.
4. Балаж А., Блажек И. Эндогенные ингибиторы клеточной пролиферации. Будапешт. М.:Мир. 1982. 302 с.
5. Биология старения. Серия "Руководство по физиологии". Академия наук СССР. Л.:Наука. 1982. 616 с.
6. Ваганов Ф.В. О роли лимфоцит-фибробластных контактов в заживлении ран // Архив патологии. 1984. N 6. С.47.
7. Девойно Л.В., Ильюченок Р.Ю. Моно-аминэргические системы в регуляции иммунных реакций. Новосибирск. Наука. 1983.
8. Донцов В.И. Влияние Т-лимфоцитов на пролиферацию клеток слюнных желез крыс и мышей под действием изопротеренола // Бюл. эксперим. биол.мед. 1985. N 7. С.65-68.
9. Донцов В.И. Системный подход к анализу процесса старения // Физиология человека..1998. Т. 24. N 1. С. 82-87.
10. Донцов В.И. Влияние лимфоцитов на рост перевиваемой аденокарциномы толстого кишечника у мышей // Экспер. онкол.1989. Т. 11. N 5. С. 48 - 51.
11. Донцов В.И. Регуляция лимфоцитами клеточной пролиферации - альтернатива теории “проти-воопухолевого надзора”? // Иммунология.
1989. N. 5. С. 94-96.
12. Долгушин И.И. Ускорение ожоговой регенерации кожи при переносе лимфоцитов от ожоговых больных // Патол. физиол. эксперим. терап. 1978. N 6. С. 30-33.
13. Донцов В.И. Регуляция индуцированной изопротеренолом гиперплазии слюнных желез молодых и старых мышей веществами, влияющими на серотонин- и дофаминэргические системы // Бюл. эперим. биол. мед.1986. N 4. С.479.
14. Донцов В.И. Применение теории гиперцикла для анализа процессов межклеточной регуляции пролиферации тканей: доказательства существования специализированной клеточной системы регуляции пролиферации тканей // Успехи современной биологии. 1986. Т. 101. Вып.1. С. 18.
15. Донцов В.И. Иммунобиология постнатального развития. М.:РАН, МОИП. Наука. 1990.
16. Донцов В.И., Крутько В.Н., Подколзин А.А. Старение: механизмы и пути преодоления. М.: 1997.
17. Долгушин И.И. Ускорение ожоговой регенерации кожи при переносе лимфоцитов от ожоговых больных // Патол. физиол. эксперим. терап. 1978. N 6. С. 30-33.
18. Зинзар С.И., Лейтина Б.И., Туманян Б.Г., Свето-Молдавский Г.Я. Злокачественные опухоли, возникающие из сингенного трансплантата эм-брионального желудочно-кишечного тракта // Вопр. онкологии 1972. Т.18. С. 89-92.
19. Крутько В.Н., Донцов В.И. Проблемы старения с позиций системного подхода //
Системные исследования. Российская академия наук. Институт системного анализа РАН. М.:1996. С.329-348.
20. Ли С.Е., Иванец Т.А., Турченко Г.В. Изменение показателей иммунитета у интактных и частично гепатэктомированных мышей при транс-плантации клеток селезенки гипокинетических доноров // Бюл. эксперим. биол. мед. 1985. N 4. С. 464.
21. Макинодан Т., Юнис Э. п/ред. Иммунология и старение. М.: Мир. 1978. 278 с.
22. Малыгин А.М., Погодина О.Н.,Чернышева М.Д., Фель В.Я. Исследование противо-опухолевого действия лимфоцитов молодых и гепатэктомированных мышей линии С3НА // Иммунология. 1984. N 4. С.46.
23. Мамонтов С.Г. Суточные ритмы митозов в эпителии рогового слоя после спленэктомии // Бюллетень экспериментальной биологии и меди-цины.1977. N 6. С. 594.
24. Мартыненко Ф.П., Шостак И.Н. Влияние соматотропина на образование аутологичных розеток тимоцитами гипотиреоидных крыс // Доклады АН УССР 1982. Серия Б. N 2. С. 67.
25. Николис Дж. Динамика иерархических систем: эволюционное представление. М.: Мир. 1989. 486 с.
26. Петров Р.В., Сеславина Л.С. Взаимодействие лимфоцитов с кроветворными стволовыми клетками // Журн. микробиол.1977. N 11. С.28-55.
27. Подколзин А.А., Донцов В.И. Старение, долголетие и биоактивация. М.: Московские учебники и картолитография.1996.
28. Подколзин А.А., Донцов В.И. Факторы малой интенсивности в биоактивации и иммунокоррекции.М.: Московские учебники и картолитография. 1995.
29. Пригожин И. Введение в термодинамику необратимых процессов. М.: Иностр. лит. 1960.
30. Свет-Молдавский Г.Я., Шхвацабая И.К., Зинзар С.Н. Изучение пассивного переноса лимфоидными клетками компенсаторной гипертрофии миокарда // Доклады АН СССР. 1974. Т.218. N.4. С. 246.
31. Тимошкевич Т.В., Харлова Г.В., Юдина Н.В. Передача регенераторной информации лимфоцитами крыс с обширной резекцией кишечника // Бюл.эксперим.биол.мед. 1984. N 3. С. 352.
32. Шилова Л.Я., Полторакин В.С., Суслова А.Г. Влияние лимфоцитов селезенки доноров, обработанных четыреххлористым углеродом, на митотическую активность печени и продукцию альфа-фетопротеина у сингенных мышей // Бюл. эксперим. биол. мед. 1982. N 6. С.99-101.
33. Эйген М., Шустер П. Гиперцикл: принципы самоорганизации макромолекул. М.: Мир. 1982. 270 с.
34. Behferanz M., Eardley D.D., Gerny J. Nonspecific immunoenhancing T cells in tumourbearing mice imclude antiidiotypic subsets / /J. Immunol. 1983.V.131.P.2576.
35. Indveri F., Barabion A., Pierri I., Grifoni V. Human autologous mixed lymphocyte reactions // Ric.Clin.e lab.1983. V. 13. P.397.
36. Horowitz M., Vignery A., Gershon R., Baron R. Thymus-derived lymphocytes and their interactions with macrophages are required for the production of osteoclast-activated factor in the mouse // Proc. Nat. Acad. Sci.USA.1984.V.81.P.2181.
37. Moody C.E.,Hausman P.B.,Gutowski J.K. etc. The developmental biology of the syngenic mixed lymphocyte reaction // Behring Inst. Mitt. 1983.N. 72. P.7.
38. Ponzio N.M. Similarities between the syngenic mixed lymphocyte response and lymphoma-induced T cell proliferation in SJL/J mice // Behring Inst. Mitt. 1983. N. 72. P. 28.
39. Schneider G.B., Relbsom M. Immunological competence in osteopetrotic rets // Immunology. 1984. V. 167. P. 318.
40. Turing A. The chemical basis of morpho-genesis // Trans. Roy. Soc. 1952. V. 237. P. 32.
Комментарии
ПРАКТИКА ПЕДИАТРА



