Сущностные модели старения и продолжительности жизни
СтатьиОпубликовано в журнале:
Профилактика старения »» Выпуск 1 1998 В.И. Донцов, В.Н. Крутько
Национальный геронтологический центр
Теоретическая биология старения и геронтология переживают кризис, связанный с тем, что старые принципы создания концетуальных моделей старения, сводившиеся по существу к абсолютизации отдельных наблюдаемых явлений и частных механизмов старения, потерпели крах. С другой стороны, ряд чисто математических подходов к моделированию старения не встречают интереса и признания среди биологов ввиду явно биологически не обоснованных и фактически не верных изначальных предпосылок. В то же время, существует настоятельная необходимость в четком общем взгляде на явление старения в целом и в моделях, позволяющих количественно и содержательно интерпретировать старение организмов, при этом можно видеть, что многие элементы таких моделей уже существует в различных областях биологии. В настоящее время важнейшим представляется достаточно подробная разработка сущностных моделей старения, отражающих само существо этого общего для всего живого явления и являющихся биологически обоснованными и биологически содержательными. Рассматриваются требования к сущностным моделям старения и ряд наиболее отвечающих этим требованиям теорий старения: теория Гомперца, теории потери информации, регуляторная теория и др, Уточняются понятие биологического возраста и максимальной длительности жизни, развивается общебиологический взгляд на природу человека и сущность старения как общебиологического явления.
1. СУЩНОСТНЫЕ МОДЕЛИ - ОСОБЕННОСТИ И ЗНАЧИМОСТЬ
Создание теоретических моделей изучаемого процесса - важнейший элемент познания, поэтому данному вопросу уделяется центральное внимание в любой современной области науки. Геронтология в этом отношении переживает кризис, связанный с тем, что старые принципы создания концептуальных моделей старения, сводившиеся по существу к абсолютизации отдельных наблюдаемых явлений и частных механизмов старения, потерпели крах. Все так называемые теории старения, которых на настоящее время насчитывают более ста, оказались несостоятельными и во многом представляют только исторический интерес.
С другой стороны, ряд чисто математических подходов к моделированию старения не встречают интереса и признания среди биологов, так как даже при самом поверхностном изучении видны биологически не обоснованные и фактически не верные изначальные предпосылки. Так, например, модные экологические и эволюционные математические теории старения, основанные на представлении о "целесообразности старения" как механизма ускоренного обновления вида, игнорируют очевидный факт высокой естественной смертности в дикой природе, когда старые животные фактически отсутствуют в популяции и практически все животные умирают молодыми.
В то же время, существует настоятельная необходимость в четком общем взгляде на явление старения в целом и в моделях, позволяющих количественно и содержательно интерпретировать старение организмов, при этом можно видеть, что многие элементы таких моделей уже существует в различных областях биологии.
В настоящее время важнейшим представляется, таким образом, достаточно подробная разработка сущностных моделей старения, отражающих само существо этого общего для всего живого явления и являющихся биологически обоснованными и биологически содержательными.
Сущностные модели должны отвечать следующим требованиям:
- иметь изначально четкое представление о биологическом содержании каждого моделируемого элемента,
- четкое представление о биологической значимости получаемых при моделировании результатов,
- четкое представление о значении модели в общем моделируемом процессе (какая часть более общего процесса описывается),
- какие иные реальные процессы происходят, принципиально влияющие на моделируемый процесс,
- существует ли моделируемый процесс реально в природе.
2. ТЕОРЕТИЧЕСКИЕ МОДЕЛИ СТАРЕНИЯ
ОБЩАЯ МАТЕМАТИЧЕСКАЯ МОДЕЛЬ СТАРЕНИЯ Б.ГОМПЕРЦА
Первая осознанная четкая математическая модель старения была создана около 200 лет тому назад Б.Гомперцом (1825). Она до сих пор наиболее точно описывает смертность человека и, видимо, большинства других организмов. В основу ее было положено очень древнее представление о потере в течение жизни общей ЖИЗНЕСПОСОБНОСТИ организма - учение о "жизненной силе" ("энтелехии" Аристотеля), которая имеет тенденцию только растрачиваться со временем для любого родившегося организма.
Являясь специалистом по страхованию жизни, Гомперц теоретически вывел практически необходимую для его профессии формулу интенсивности смертности, повышение которой во времени и служит наиболее общим определением старения как такового до настоящего времени.
Смертность, как количественную характеристику "неспособности противостоять разрушению", Гомперц рассматривал как величину, обратную жизнеспособности - способности противостоять всей совокупности разрушительных процессов. Он предположил, что во времени жизнеспособность снижается пропорционально ей самой в каждый момент, что для смертности соответствует экспоненциальному нарастанию с возрастом.
Такое неспецифическое повышение уязвимости организма ко всем воздействиям с возрастом и носит название старения как такового. Сам подход к написанию формулы в настоящее время теоретически понятен: это элементарное дифференциальное уравнение, описывающее, например, радиоактивный распад в физике и иные простые вероятностные процессы; сущность процесса в том, что в каждый момент времени изменение состояния не зависит от предыстории, а только от настоящего состояния системы.
Понятны и общие механизмы таких процессов - это принципиально вероятностные закономерности, связанные с конечной устойчивостью любых отграниченных реально существующих неизменных элементов; тогда сложный организм, состоящий из таких элементарных единиц, может со временем их только утрачивать. Главным вопросом является в таком случае природа таких "элементарных единиц жизни".
Гомперц отмечал сходство кривых изменения смертности и энтропии, а В.Перкс (1932) писал (16) , что "неспособность противостоять разрушению имеет ту же природу, что и рассеяние энергии" (то-есть, старение эквивалентно увеличению энтропии, которая служит мерой неупорядоченности любой системы). А.Комфорт в "Биологии старения" (1967) пишет о том, что "жизненность" на современном уровне понимания может быть сведена к достаточно конкретному, хотя и не вещественному субстрату - "в настоящее время представляется вполне вероятным, что информация, содержащаяся в клетках, и есть... "биологическая энергия".
Таким образом, содержательная интерпретация понятия "жизнеспособности" с самого начала сводилась и сводится в настоящее время не столько к вещественному наполнению ("энтелехия" древних), сколько к энергетическому и информационному содержанию.
В соответствии с разрабатываемой Б.Гомперцом моделью жизнеспособность (А) снижается во времени пропорционально ей самой (Рисунок 1): dA/dt = - k A, где k - коэффициент.
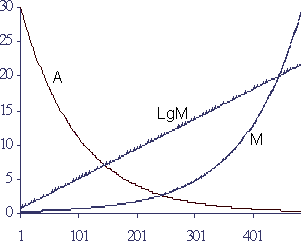
Рисунок.1
Моделирование потери жизнеспособности и нарастания смертности по формуле Гомперца
По вертикали - значения параметров в условных единицах, по горизонтали время в условных единицах. А - жизнеспособность (при исходной Ао = 30 условных единиц и 1% гибели за единицу времени), М - смертность (М=(1/А)7), LgM - логарифм смертности (Lg(5М)10).
Смертность (М), как противоположность жизнеспособности, можно определить как М=к 1/А, где "к"- коэффициент пропорциональности. Тогда повышение вероятности смертности для отдельного организма (или, что то же самое, повышение доли умерших в популяции) во времени будет происходить по экспоненте, а логарифм "М" будет меняться по линейному закону, что и наблюдается в действительности - конечная форма уравнения Гомперца принята: M(t)=Ro exp(a t),
где "М" - смертность, изменяющаяся во времени - "t"; "Ro" - начальный уровень смертности, "а" - коэффициент, характеризующий скорость нарастания смертности со временем).
Эта формула была впоследствии модифицирована затем У.Мейкемом, добавившим в формулу Гомперца постоянный коэффициент, представляющий независимый от возраста компонент смертности, имеющий, как теперь становится ясно, эколого-социальную природу и выраженно меняющийся в истории человечества: М(t) = A + Ro exp(a t)
До настоящего времени формула Гомперца-Мейкема остается наилучшей для описания смертности, связанной со старением, для самых раз-личных видов, включая человека.
Принято представлять график экспоненты в полулогарифмических координатах, где она имеет вид прямой. Однако, аддитивная поправка Мейкема в правой части уравнения обусловливает отклонение от прямой линии зависимости Ln(m) от t. Поскольку А является константой, получить в правой части уравнения чистую экспоненту можно, продифференцировав уравнение.
Для целей количественной геронтологии необходимо иметь возможность вычисления параметров уравнения Гомперца-Мейкема, что можно сделать методами нелинейной регрессии. Для вычисления "вручную" Л.А. Гаврилов и Н.С. Гаврилова (1991) предлагают нижеследующий алгоритм, дающий вполне удовлетворительную точность (1). Пусть мы имеем значения чисел доживших "l" для четырех равноотстоящих друг от друга моментов времени: t, t+n, t+2n...
Вначале вычисляют вспомогательные величины:
y1 = ln[ l(t)/l(t+n)];
y2 = ln[l(t+n)/(l(t+2n)];
y3 = ln[l(t+2n)/(l(t+3n)];
z = y1+y3-2y2;
w= (y3-y2)/(y2-y1).
Тогда параметры формулы Гомперца-Мейкема могут быть найдены из следующих соотношений:
A = (y1 y3-Y22)/(z n),
Ro = [(y2-y1)2ln(w)]/[z n(w-1)wt/n],
a = ln(w)/n,
Можно продолжить эти рассуждения и получить, например, закон изменения выживаемости популяции (для начального "N" ее членов) с возрастом: доля умерших будет во времени пропорциональна в каждом возрастном интервале величине смертности для этого интервала: N(t) - N(t+1) = k M N(t), что дает хорошую аналогию с реальными кривыми выживаемости и смертности (Рисунок 2).
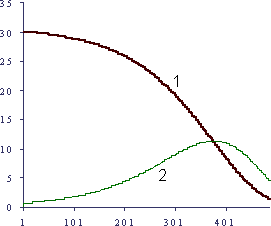
Рисунок. 2
График расчетной выживаемости когорты для модели старения по Гомперцу
По вертикали - значения параметров в условных единицах,по горизонтали время в условных единицах. 1 - выживаемость ( N(t) для начальной No=30), 2 - умершие (X(t)-X(t+1) )100. М – как для рисунка 1, k принято равным 0,001.
Интересны некоторые очевидные и экспериментально известные выводы, иногда, однако, парадоксально звучащие. Так, например, очевидно, что наибольшее абсолютное снижение жизнеспособности можно наблюдать в раннем возрасте, что мы и видим по кривым изменения в онтогенезе абсолютного значения многих физиологических функций. В это время, соответственно, эффективны мероприятия по профилактике старения и удобно проводить экспериментальную проверку геропрофилактических средств. В то же время, в старости даже небольшие абсолютные изменения жизнеспособности ведут к выраженным изменениям смертности, поэтому в старших возрастах удобно изучать влияния адаптогенов и биостимуляторов, хотя малый жизненный ресурс может и не приводить к повышению длитель-ность жизни при их использовании.
Полученная кривая изменения жизнеcпособности, однако, все же отличается от реальной в ее начальной и конечной части и отражает изменение уже сформировавшегося организма, прекратившего процессы роста и развития.
Кроме того, указанный механизм по существу игнорирует качественное отличие живого и неживого вещества - способность первого из них к самообновлению (путем самокопирования) и самоорганизации, причем последняя проявляется как спонтанная тяга к устойчивости при выводе сложной системы-организма из равновесия в ходе жизнедеятельности, а также в виде процессов роста и развития в течение жизни. Можно, однако, достаточно просто промоделировать (для наших целей) и эти особенности живого организма в наиболее общем виде. При этом следует учитывать основное теоретическое положение, что самообновление как спонтанный процесс (метаболизм, клеточная пролиферация) имеет несопоставимо высокие временные масштабы в сравнении с очень длительно протекающими процессами старения и развития (микросекунды для метаболизма, часы и сутки для клеточного деления и десятки лет для роста и развития). Поэтому длительные изменения самообновления во времени возможно только как изменения фоновых внешних влияний на них - по-существу, как регуляторные влияния.
РЕГУЛЯТОРНАЯ МОДЕЛЬ СТАРЕНИЯ
Центральным для жизни как таковой является процесс постоянного самообновления всех составных частей организма, что, по существу, тождественно определению жизни как таковой. Вполне логично представить, что уровень самообновления определяет и уровень жизнеспособности. Однако, высокие скорости процессов самообновления для уровня биохимических процессов (мкСек) и даже для клеточных процессов (часы) однозначно указывают, что на этих уровнях самообновление должно стремиться, что на самом деле и имеет место, к стабилизации в каждый момент времени. Действительно, с точки зрения современных теорий самоорганизации и кибернетики биохимические процессы и клеточное самообновление - это саморегулирующиеся и самостабилизирующиеся системы. Для них нет никаких процессов, которые способны были бы во времени постоянно и однонаправленно изменять состояние всего организма в одном направлении - процессов развития и старения.
Для осуществления долговременных процессов развития организм использует специальные уже чисто регуляторные, механизмы, функционирующие на уровне целостного организма. В ряде случаев осуществление таких регуляторных программ может приводить к снижению самообновления, а, значит, и жизнеспособностии - к старению (Давыдовский, 1966, Донцов, Крутько, Подколзин, 1997, Подколзин, Донцов, 1997, Фролькис, 1975).
Рассмотрим следующую простую схему. Пусть уровень самообновления (например, уровень клеточного самообновления путем деления клеток в популяции, ограниченной и стабилизированной кейлонными и иными обратными связями) будет пропорционален некоторому веществу "С". Вследствие быстрого установления нового равновесия для делящейся клеточной популяции при повышении содержания регуляторного вещества в окружающей клетки среде и необходимости сохранить длительно рост всего организма (повышение массы с 4-6 кг при рождении до 50-80 кг за 20 лет), в течение всего периода роста организма нужно сохранять повышающийся градиент вещества "С", например, в крови. Это можно обеспечить, например, за счет растормаживания вырабатывающих это вещество регуляторных "s" клеток. Реально такие процессы широко известны, например, для нейрогормональной регуляции и осуществляются за счет спонтанной высокой гибели клеток-ингибиторов "i" в регуляторных центрах вегетативного мозга (гипоталамуса).
При наличии различий в спонтанной гибели клеток стимулирующего "s" и "i" типов будет иметь место повышение уровня стимулирующего фактора "F" некими регуляторными клетками следующего иерархического уровня организации (уровень "F" будет пропорционален разности "s - i"), что обеспечивает рост организма за счет высокого уровня клеточной пролиферации, а значит, и клеточного самообновления, что важно, например, для кожи, слизистых, печени, почек и других органов с высоким уровнем клеточного самообновления.
Спонтанное снижение количества "s" и "i" можно описать обычным образом: ds/dt = - k1 s; di/dt = - k2 i,
где k1 и k2 - коэффициенты, причем по нашему условию k2 > k1.
В то же время, наличие спонтанной гибели не только "i", но и "s" клеток, что типично для необновляющихся нервных клеток, приведет после достижения пика к снижению уровня "С" фактора и к снижению интенсивности самообновления. Приняв, что жизнеспособность организма пропорциональна самообновлению, а смертность - обратно пропорциональна ему: М=1/(F+с),
где "с" отражает некоторый изначальный уровень устойчивости в отсутствие регуляторного фактора, мы можем видеть (Рис. 4 и 5), что кривая, отражающая изменения смертности, полностью моделирует все особенности изменения смертности организма с возрастом: высокую начальную смертность, связанную с малой жизнеспособностью еще не сформировавшегося организма, экспоненциальное повышение уровня смертности после окончания развития и конечный "загиб" ее в самых старых возрастах, когда регуляторные влияния уже не играют особой роли в общей устойчивости организма, зависящей теперь уже от более иерархически "глубоких" слоев его организации.
Таким образом, регуляторная теория старения оказывается достаточно простой и эффективной при качественном и количественном описании процессов старения реальных систем. Более того, она едва ли не единственная может объяснить причины высоких видовых различий по срокам жизни для близких видов, построенных из сходных типов клеток и тканей (например, для человека и мыши) и возможности в ряде случаев резко, в разы, а для нетеплокровных и на порядки, изменять длительность жизни организмов в эксперименте или при изменении естественных условий. Однако, для каждого конкретного организма применение регуляторных теорий требует ответа как минимум на следующие вопросы:
а) какую часть от общего процесса старения занимает регуляторный процесс;
б) на что конкретно он влияет и каковы конкретные механизмы его реализации;
в) какие симптомы старения тканей отвечают в первую очередь действию данного механизма, а какие - вторичны и определяются другими механизмами старения,
г) как данный общий механизм старения взаимодействует с другими типами старения, насколько обратимы вызываемые эффекты старения клеток и тканей,
д) каковы реальные возможности повлиять на этот процесс для данного организма и в каких пределах;
е) как будет выглядеть процесс старения данного организма в отсутствие регуляторного компонента процесса старения у него.
Совершенно ясно, например, что для мышей, у которых имеет место различие в разы длительности жизни весенней и осенней генераций потомства в естественных условиях регуляторные механизмы принципиально более важны, чем для че-ловека, у которых фактически нет сезонных ритмов старения. Ясно также и то, что изменения тканей 2-х летней мыши, аналогичные изменениям однотипных тканей 60-80-летнего человека в принципе также в большой мере зависят от регуляторных влияний, тогда как живущий в течение 100 лет человек гораздо менее подвержен, видимо, действию собственно регуляторных механизмов.
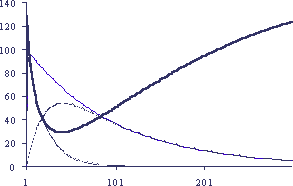
Рисунок.4
Изменение смертности при регуляторном ограничении самообновления
По вертикали - приведенные значения параметров, по горизонтали - время в условных единицах. 1 - количество клеток-стимуляторов (s) для на-чального s = 100 при спонтанной гибели 1% клеток за единицу времени, 2 - количество клеток-ингибиторов (i) для начального i=100 при спонтанной гибели 5% клеток за единицу времени, 3 - содержание регуляторного фактора (F = s - i), 4 - логарифм смертности (LgМ=Lg(1/(F+с)100)150) при с=10)
НОВАЯ ИММУННАЯ ТЕОРИЯ СТАРЕНИЯ
Оригинальными российскими исследованиями, в том числе нами показано, что в современном многоклеточном организме существует специальная система регуляции клеточного роста любых соматических пролиферирующих клеток, представленная, в частности, субпопуляциями Т-лимфоцитов (система КРП).
Более подробно этот вопрос рассмотрен в отдельных публикациях (Донцов, 1990, Донцов, Крутько, Подколзин, 1997, Подколзин, Донцов, 1997) и в отдельном разделе книги.
Показано рядом авторов, что эта система непосредственно связана и с регуляцией роста целостного организма, с процессами регенерации, гиперплазии, опухолевого роста и со снижением уровня клеточного самообновления в старости, когда снижается скорость клеточного деления самых различных типов соматических клеток.
Нами была выдвинута новая иммунная (лимфоидная) теория старения, связывающая возрастное снижение клеточного роста и самообновления непосредственно с регуляторными изменениями в Т-лимфоидной системе иммунитета, в той ее части, которая регулирует клеточный рост соматических клеток (Донцов, 1990-2000).
Может показаться, что хорошее соответствие полученных результатов для "регуляторной теории" старения позволяет быстро и просто обратить старение внешним введением регуляторного фактора "F. Однако, в реальности в организме протекают процессы старения и не связанные с самообновлением, представленные выше и отражающие, например, необратимую гибель нервных клеток, альвеол, нефронов, отдельных уникальных генов в клетке, даже зубов и пр. Именно этим механизмам принадлежит, видимо, центральная роль в старении рыб и иных постоянно растущих в течение всей жизни организмов.
Фактически, в организме имеется огромное число необновляющихся на своем иерархическом уровне элементов, стареющих необратимо по механизму простой утраты клеток. Не трудно видеть, что этот же механизм лежить в основе и разобранных регуляторных механизмов - так стареют нервные регуляторные центры. Вклад данного механизма в старение достаточно высок - к концу жизни, например, человек теряет более половины нефронов в почках, альвеол в легких, более 70% клеток некоторых регуляторных центров гипоталамуса и пр. Сформировавшиеся вторичные изменения, например, компенсаторное увеличение объема альвеол и возрастная эмфизема, не обратимы. Вот почему все методы биостимуляции как правило не приводили к выраженному повышению продолжительности жизни - старый организм не способен поддерживать высокий уровень самообновления и функционирования, поэтому снижение самообновления в старости - и механизм старения и компенсационный процесс для старого организма.
СТАРЕНИЕ КАК СПОНТАННАЯ ПОТЕРЯ И ИЗМЕНЕНИЕ ИНФОРМАЦИИ
Существуют еще минимум два механизма старения, вносящие свой вклад в старение целостного организма.
Это, во-первых, изменение при старении обмена веществ (а также энергии и информации) с внешней средой и, во-вторых, повышение степени разнообразия для самых разных структурных элементов и связей в организме - "разрегулирование" целостной системы организма.
Оба механизма являются конкретизацией процесса спонтанной утраты информации в системе, за которым следует ее материальная и энергетическая деградация. Действительно, положим, что некоторая система (организм) получает извне поток вещества (Р1), энергии (W1) и информации (I1). Сохранение самое себя системой означает поддерживание постоянства материальной струк-туры системы (p2), энергетических потоков и взаимосвязей (w2) и тождества во времени информации о себе (i2); заметим также, что, так как организм только часть большего его некоторого целого - биосферы, например, то (P1, W1, I1) >>> (p2, w2, i2).
Кроме того, сохранение системы во времени означает тождество суммарных потоков, поступающих из внешней среды, выводящейся из системы и сохраняющегося динамически потока внутри системы. Не трудно видеть, что центральным при таком рассмотрении оказывается процесс сохранения информации в системе, т.к. вещественная и энергетическая организация являются только "материальными носителями" этой информации и, фактически, следуют качественно и количественно за изменением информации, которая выступает как регулирующий, управляющий и (само)-организующий фактор.
В общем виде информация в системе может изменяться благодаря следующим процессам:
- поступлению информации (и энтропии) извне (например, "ремонт" силами извне или эволюционное давление при формировании новых признаков и т.п.);
- появление новой информации (и энтропии) внутри системы за счет взаимодействия в ней вещества и энергии в ходе сложнейших взаимопревращений и взаимосвязей (метаболизм, рост и развитие, механизмы саморегуляции и самоорганизации и т.п.);
- изменение и потеря информации в системе (развертывание программ роста и развития; "мутации" материальных носителей информации - ДНК, белков и иерархически других структурных уровней материальных носителей информации; спонтанный распад информации - производство энтропии и т.п.).
Процесс утраты информации аналогичен ее изменению - "мутациям", причем он носит вероятностный характер и, по существу, сводится или к ошибкам в ходе процесса воспроизводства информации в ходе самокопирования материальных носителей информации, или к спотанному вероятностному "мутированию" невоспроизводящейся информации (например, повреждения свободными радикалами неделящейся ДНК и т.п.). Заметим, что во многих случаях "мутировавшая" информация способна к воспроизводству (например, большинство мутаций клеток не приводят к прекращению их деления) и часто сохраняется возможность функционирования воспроизводящихся на ее основе структур, которые, таким образом, вступают в конкуренцию с имеющимися ранее структурами организма.
Исходя из выше сказанного, можно свести главные процессы воспроизводства и изменения информации (и ее материально-энергетических носителей) в организме к следующим уравнениям. Для точного воспроизведения информации (I) используется механизм самокопирования, который сопровождается ошибками с воспроизведением измененной (Im) и некоторой необратимой потерей информации (анаболизм, распад и полная деградация структур с выведением вещества и энергии их из организма и рассеиванием их информации): dI/dt = k1 I - k2 Im - k3 I
Соответственно, "мутировавшая" информация может также воспроизводиться сама, пополняться за счет мутаций неизменившейся информации и рассеиваться (обратная мутация в исходную форму крайне мало вероятна и очень мала): dIm/dt = k4 Im + k2 Im - k5 Im
Для того, чтобы учесть требуемое сохранение постоянства вещества, энергии и информации после прекращения развития у взрослого организма, введем в формулы ограничение количества информации (I+Im=const), получив известную из кибернетики формулу самовоспроизводящейся системы с обратными положительными и отрицательными связями: dI/dt = k1 I / k4 (I + Im) - k2 Im - k3 I; dIm/dt = k5 Im / k4 (I + Im) + k2 Im - k6Im
Численная модель рассеивания начальной ин-формации в стабилизировавшейся системе представлена на рисунке 5.
При анализе модели учтено, что мутировавшие клетки обычно менее жизнеспособны и, кроме того, подвергаются иммунному надзору и гибнут поэтому быстрее, а также по тем же причинам с меньшей скоростью самообновляются. Соответственно коэффициенты для модели подобраны в случае графика: k1=0,3, k5=0,2, k4=0,1, k2=0,03, k3=0,05, k6=0,07.
На модели можно видеть, что со временем соотношение мутантных и неизмененных единиц информации стабилизируется, но в течение некоторого периода будет иметь место нарастание числа мутаци, что будет вести к нарастанию смертности. Вид кривой смертности, однако, не экспоненциальный, а линейный, а лоарифма смертности – выпуклый, что значительно отличается от реальной кратины. Это не удивительно, так как время установления равновесия I и Im невелико – фактически, например, время клеточного деления для клеток слизистой и кожи – дни и часы, поэтому на фоне многих лет жизни напрямую этот механизм вряд ли вносит существенный вклад в процесс старения. Накопление мутаций скорее отражает другие процессы – резкое (регуляторное) снижение скорости клеточного самообновления и снижение эффективности иммунного надзора с возрастом. Мутации важны и в случае повышения с возрастом риска возникновения опухолей, что вносит значительный вклад в причины смертности для млекопитающих вообще и человека в особенности.
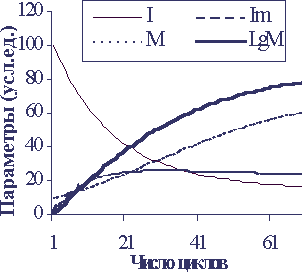
Рисунок 5
Спонтанное рассеивание информации в сложных обновляющихся и мутирующих системах
По вертикали - значения параметров в условных единицах, по горизонтали - время в условных единицах.
I - количество начальной информации, Im - количество измененной информации, M - смертность, LgM - логарифм смертности. Коэффициенты подобраны для удоства отображения графика.
Так как мутации возможны самые разнообразные, то фактически за счет этого же механизма мы имеем и второе характерное для старения следствие: увеличение разнообразия исходно однородных структур. Увеличение разнообразия структур - появление большого количества "чужой" информации перегружает системы организма, ответственные за распознавание и удаление ее, причем, т.к. фактически, новые структуры лишь немного отличаются от старых и сохраняют во многих случаях практически на прежнем уровне функциональную способность и, соответственно, реальную ценность для организма, то чрезмерная реакция против "чужого" даже вредна.
Это известно на примере повышения уровня аутоантител против собственных структур организма с возрастом (результат реакции иммунной системы на "изменившееся свое") и снижении длительности жизни при слишком высоких уровнях таких антител. Фактически, организм со временем выходит на разумный балланс между аутоиммунным саморазрушением и неконтролируемым, в том числе опухолевым, ростом и разнообразием, причем оба процесса в конечном счете разрушительны.
Кроме того, так как информация контролирует потоки вещества и энергии, входящие в систему и выходящие из нее, то изменение балланса I/Im фактически означает снижение возможности отбора "нужной" вещественной и энергетической основы для строительства своей структуры и снижение распознавания и вывода "чужого" вещества и энергии - то-есть, отражает известный механизм старения - "накопления шлаков": ( d(I/Im)/dt = d(p2,w2/P1,W1)/dt ).
Полные математические выкладки для иллюстрации выше сказанного достаточно трудоемки, однако, не трудно видеть, что в общем виде оба процесса старения, как впрочем и уже описанные выше, это только частные проявления процесса нарастания энтропии, рассматриваемые с различных "точек зрения" - с точки зрения различных свойств живого вещества, живых систем. Действительно, и спонтанная потеря жизнеспособности в целом, и регуляторное снижение самообновле-ния, и снижение структурной однородности и "загрязнение" организма не выведшимися и отложившимися "балластом" веществами - все это в глобальном плане есть отражение действия единственной причины - дискретности организма, действия законов термодинамики на частично открытую систему, не способную, после окончания развития, к эффективной дальнейшей эволюции. Заметим, при этом, что старение не есть "выработавшийся в эволюции феномен", нужной для исключения неэффективных форм жизни и смены новыми, старение отражает более глобальные закономерности Бытия вообще.
ОБЩИЕ ЧЕРТЫ ИЗМЕНЕНИЯ МОРФОЛОГИИ И ФУНКЦИИ СТРУКТУР И ПРОЦЕССОВ В ХОДЕ СТАРЕНИЯ
Общее определение процесса старения позволяет, в частности, описать общие черты возрастной эволюции морфологии и функции любых структур и процессов в организмах любых видов.
Во времени нарастает разнообразие структуры и функции и общая энтропия - "степень накопления хаоса". Иерархичность всех структур (органы-ткани-клетки-молекулы) определяет участие многих однородных элементов в каждом процессе, а разнообразие их приводит к случайному закону распределения для любых регистрируемых параметров морфологии и функции. Это проявляется как "размывание" любых характеристик со временем, причем наблюдается общая тенденция (сдвиг) в сторону ухудшения.
БИОХИМИЧЕСКИЕ И ТЕРМОДИНАМИЧЕСКИЕ МОДЕЛИ СТАРЕНИЯ
Этот уровень рассмотрения позволяет понять что стоит за "жизнеспособностью", приписываемой не только всему организму, но и каждому его элементу. Действительно, "старой" является каждая часть старого организма, но тогда возникает вопрос - какая характеристика, общая для самых разнообразных структурных элементов организма, может отражать эту жизнеспособность и величину, ей обратную - уровень старения, для самих молекул и их комплексов, химических реакций и т.п.
С точки зрения биохимии и теорий сложных систем на такую характеристику претендуют две (тесно взаимосвязанных) общие характеристики, применяемые в химии, биохимии и вообще теории сложных систем: это энтропия и информация (2, 3, 5, 9-11, 13, 15-18). При этом энтропия (S) может быть прямо определена через информацию, которая, в свою очередь, тесно связана с вероятностью события: S = A LnW + B,
где А и В - коэффициенты, а W - вероятность события.
Уже из этого видно, что максимум энтропии соответствует максимально вероятному состоянию, поэтому жизнь с точки зрения термодинамики определяют как степень отклонения от максимально вероятного состояния - от равновесия; как "неравновесный поток", как отклонение от состояния равновесия.
Соответственно, можно теперь представить уровень жизнеспособности как степень отклонения от состояния равновесия.
Очевидным при таком рассмотрении является и то, что для поддержания жизнеспособности в любой части любой живой системы необходима постоянная затрата энергии, так как для живых систем характерно снижение энтропии, а согласно второму закону термодинамики это возможно только при постоянном поступлении энергии извне.
Таким образом, по степени потребления энергии организмом можно судить о степени его жизнеспособности.
На практике используют оценку общего обмена для целого организма, скорость потребления кислорода тканями, уровень потребления кислорода при нагрузке ("коэффициент полезного действия" и мощность живых процессов), а также максимально возможное потребление кислорода и/или мощность внешнего развиваемого усилия - объем адаптации организма. Предлагаются также и некоторые иные подходы. Например, связь энтропии с теплосодержанием позволяет, в принципе, по микро-калориметрии нативной и денатурированной ДНК говорить в энергетическом эквиваленте о том, какая энергия и, соответственно, энтропия, соответствует "живому" и "неживому" состоянию даже для отдельной молекулы. Однако, интерпретации такого рода с привлечением физических эквивалентов живого, достаточно трудны.
Для целостного организма эффективными, видимо, являются подходы оценки энтропии с точки зрения информации и ее гармоничности для целого единого организма. Исходят из представлений об оптимальности взаимосвязей различных регуляторных систем: в каждый момент времени возможно оптимальное гармоничное состояние, обеспечивающее максимальную жизнеспособность, максимальную адаптацию и т.п. Исходя из кросс-корреляций между различными системами (сердечно-сосудистой, дыхательной, эндокринной и пр.) можно выразить в безразмерных числах такие коэффициенты взаимодействия и придать им значимость информационного содержания, а также выразить это в понятиях энтропии.
Сколь-либо развернутые исследования в этом направлении, однако, отсутствуют, что затрудняет практическую реализацию и интерпретацию таких подходов. Несомненно, однако, что именно энтропия является важнейшим показателем старения и способы, позволяющие вычислять ее для целостного организма должны открыть принципиально новые возможности для анализа в биологии старения и для выяснения первичных, сущностных, механизмов и первопричины старения.
Для практических целей важно рассмотреть возможности влияния на информационно-энтропийные процессы в целом, что позволило бы влиять и на сущностный механизм старения. Для разных уровней организации, очевидно, существуют разные возможности. Так, для целостного организма это уже обсуждалось в связи с регуляторными моделями старения. Возможности здесь сводяться к:
а) влиянию на процессы роста, развития и самообновления клеток (эндокринные влияния, нервные влияния и гуморальные влияния),
б) гармонизацию имеющихся процессов (акупунктура, физическая и психологическая тренировка и т.п.),
в) социально-общественные мероприятия как "здоровый образ жизни".
Особый интерес представляет возможность влияния непосредственно на химические процессы, используя термодинамические подходы. Вообще говоря, химическая смесь реально подвержена огромному числу реакций, идущих в самых разных направлениях. Жизнеспособность здесь эквивалентна выделению из химического "шума" биохимически значимой информации - возникновения направленности биохимических потоков. Наиболее интересным здесь, видимо, является рассмотрение ферментного механизма. С точки зрения биохимии и термодинамики жизнеспособность можно характеризовать как отношение скоростей ферментозависимых реакций "жизни" - информационно ценных биохомических процессов, к неферментным реакциям "шума" - энтропии. Важно, что такие процессы в целом зависят от внешних факторов - температуры, рН, окислительно-восстановительного потенциала, давления газов и пр. Интересно, что все перечисленное предлагается с успехом для биостимуляции и борьбы со старением:
а) коррекция рН, прежде всего диетотерапией - способствование ощелачиванию внутренних сред на вегетарианских режимах,
б) коррекция ОВП с использованием химическиактивированных сред, электролизеров и методов электроочистки воды, что позволяет изменять ОВП водопроводной воды, обычно имеющей ОВП порядка +600, до 0 и даже отрицательных значений; попутно меняется вторичная структура воды - она становится "гомогенной",
в) изменение давления газов широко известно как высокогорная терапия, а также реализовано в современных установках гипокси-терапии и т.п.,
г) снижением температуры объясняют многие эффекты продления жизни, как у холоднокровных (наиболее показательно), так и у млекопитающих - например, эффекты дозированного голодания у крыс обычно сопровождаются и выраженным снижением температуры тела. Длительная жизнь и эффекты омоложения, наблюдаемые у животных после зимней спячки также могут быть связаны с этими влияниями.
Наконец, в последнее время популярными стали прямые энерго-активационные методы, которые могут прямо влиять на уровень биохимических реакций, в частности, на энергии активации таких реакций, что все более широко применяется для общей биостимуляции и оздоровления:
а) лазерное облучение в т.ч. внутривенное,
б) озонотерапия,
в) электромагнитное облучение в т.ч. КВЧ-терапия и многие методы физиотерапии.
Эффекты температуры на химические процессы интересно обсудить с точки зрения влияния на соотношение фермент-зависимых и неферментных процессов. Так как оба типа процессов различаются в своей чувствительности к температуре, вообще говоря, следует ожидать для двух разных температур различия в отношении скоростей фермент-зависимых и независимых процессов, что можно выразить и в терминах энтропии. Такие подходы можно без труда реализовать в эксперименте и попробовать найти оптимальную область, где старение будет минимально. Однако, если для холоднокровных это, видимо, не важно, для млекопитающих это может представлять собой проблему. Для высших животных и человека теплокровность - важное эволюционное приобретение, позволяющее, в частности, поддерживать важнейшие новые функции - прежде всего уровень психических процессов. Поэтому реализации на практике биохимических подходов к воздействию на старение может препятствовать для человека не технология, а совсем другое - его психология, социальная сфера, мораль и другие чисто человеческие качества.
Вообще говоря, исходя из иерархичности ценностных приоритетов для человека старение все влияния на него, тем более затрагивающие саму биологическую природу человека, невозможно рассматривать без учета чисто человеческих факторов.
МНОГОУРОВНЕВЫЕ ПРОГРАММЫ РАЗВИТИЯ - ОСНОВА ЖИЗНИ
Понимание действительных целей и задач в биологии старения и геронтологии должно вытекать из реальной жизнедеятельности организма со всеми его особенностями. Выхватывание одного момента из жизни конкретного организма является типичной методологической ошибкой, повторяемой неоднократно геронтологами и биологами старения, что и приводит к типичным результатам - создаваемые теории старения все оказались частными случаями описания отельных проявлений его и не способны решить и даже поставить проблему в целом.
Главным определяющим моментом рассмотрения должен быть общебиологический подход к проблеме, рассматривающий жизнь со всеми ее атрибутами в целом.
Оказывается, такой подход известен был всегда и анализ на таком уровне проблемы дает вполне однозначные результаты, сведенные для удобства в приведенную схему.
Жизнь индивидуального организма - стабилизированный векторный (имеющий итоговое направление изменения) поток, изменяющий свои характеристики со временем. Онтогенез - процесс в развитии жизни в целом и противодействие старению, всегда подразумевает изменение темпа и направления индивидуального развития, а также продолжение всех отдельных программ развития и общего их результата для организма в целом, а не только противодействие разрушительным процессам в организме - собственно "старческим" изменениям.
Отметим то важнейшее обстоятельство, что понятие "человек" включает как минимум 3 качественно разнородных уровня и все они являются существенными для осознания себя полноценным существом - четвертый уровень, уровень самоосознания организма как единого целого:
- материальный субстрат - физико-химический уровень живого;
- чувственная оценка действительности - чувственно-эмоциально-оценочный уровень;
- мыслительно-интеллектуальный анализ действительности - способ преобразования мира;
- уровень целостности всего организма, гармонизации всех его составляющих и взаимодействий с внешней средой.
- как будет развиваться далее человечество, выйдя из-под непосредственной регуляции биосферы;
- как изменяется вид человека в целом;
- как изменится биологический носитель человека как организма;
- как изменятся чувства и интеллект человека;
- для чего, собственно, необходимы такие изменения (цель и задачи самоосознания существования человека как вида во Вселенной);
- моральные, социальные и иные вытекающие из этого проблемы, которые чем далее, тем больше запрещают возможные, но не адекватные, не отвечающие "человечности" воздействия человека на самого себя.
Реальная задача можеть звучать только так: "поставить под осознанный контроль всю программу развития человека как индивида и вида и овладеть осознанным процессом формообразования в широких пределах организма в целом".
Не следует также забывать, что человек существует и как индивид, развивающийся в онтогенезе, а как единица вида – задачи преодоления старения в том и другом случае различны.
Глобальность задачи противодействия старению - главный вывод, без учета которого истинные задачи, вытекающие из истинного состояния проблемы, подменяются ложными и частными, вытекающими как правило из личных эмоциональных влечений и узких представлений отдельных исследователей. Старение - комплексный глобальный феномен и влияния на него - глобальная задача.
ВИДОВОЙ ИЛИ "МАКСИМАЛЬНЫЙ ПРЕДЕЛ" ЖИЗНИ
Представление о максимальной (видовой) продолжительности жизни (МПЖ или ВПЖ), с которым связаны определения предела жизни, является относительно более важным моментом для геронтологии, так как средняя продолжительность жизни есть предмет изучения скорее демографии и медицины.
Исследования с помощью строгих математических выкладок позволяют в настоящее время говорить как о доказанных фактах о ряде важнейших закономерностей, характерных для биологии старения и продолжительности жизни.
К важному открытию привел анализ данных о смертности людей в различные исторические эпохи. Было показано для одного и того же региона, что если постоянная "А" в формуле Гомперца-Мейкема, отражающая условия окружающей среды, менялась значимо, то остальные параметры фактически оставались неизменными - "ФЕНОМЕН ИСТОРИЧЕСКОЙ СТАБИЛЬНОСТИ ВОЗРАСТНОЙ КОМПОНЕНТЫ СМЕРТНОСТИ", - то-есть, старение человека фактически всегда происходило одинаково и определялось практически исключительно внутренними биологическими особенностями его организма, а не характеристиками социальной среды и образа жизни.
Этот, казалось бы важный, аргумент в пользу представлений о существовании "ВПЖ", однако, имеет контраргумент, заключающийся в том, что для РАЗНЫХ регионов существуют РАЗНЫЕ показатели возрастной компоненты смертности, хотя и стабильные для ДАННОГО региона. Эти исследования открыли новое научное направление - популяционную геронтологию, предмет которой - изучение исторических, экологических, социо-культурных и генетических влияний на ВПЖ человека, а вернее, на его возрастную компоненту смертности.
Было показано, как и следовало ожидать, что компонента "А" отражает историко-социо-культурные влияния и для развитых стран в настоящее время практически приближается к нулю, что и проявляется в типичных близким к прямоугольным кривых дожития населения этих стран. Таким образом, повышение уровня и комфортности жизни, высокий уровень медицины, оптимальное питание и прочие сходные факторы развитой культурной среды практически исчерпали к настоящему времени свое влияние на ПЖ человека. Поэтому, перспективными являются исследования возможностей влиянии на его биологическую основу, определяющую возрастную компоненту смертности, изменяющуюся по экспоненте.
Исследования возрастной компоненты смертности оказали важное методологическое влияние на геронтологию, так как подтвердили общий тезис об условности и неполноте односторонних взглядов на причины старения. Так, подверглись пересмотру представления о только "внешней" или только "внутренней" природе старения, о генетических, эволюционных и экологических влияниях, влиянии традиций питания на ПЖ и др.
Было, например, показано, что практически во всех случаях показатели смертности для мужчин выраженно выше, чем для женщин (хотя в общем случае для других видов может наблюдаться как подобная, так и обратная картина).
Было найдено, например, что Финляндия характеризуется наиболее высоким уровнем биологической компоненты смертности мужчин среди европейских стран, и это сопряжено с высокими средними показателями концентрации холестерина в крови. Была показана связь этого явления с недостатком селена в окружающей среде и разработана программа профилактики. Селен, как известно, входит в активный центр ферментов, содержащих серу и участвующих в дезактивации перекисных соединений. Известно, что в "Х" - хромосоме женщин расположен ген глюкозо-6-фосфат-дегидрогеназы - ключевого фермента в образовании НАДФН. Последний кофермент важен для поддержания в восстановленной форме глютатион-пероксидазы, в состав которой как раз и входит селен. Фермент определяет защиту организма от перекисей, накопление которых - главный механизм метаболических повреждений клетки.
С другой стороны, в Дании зарегистрирована необычно высокая смертность среди женщин, что связано с традиционным избытком жирной пищи и высоким уровнем опухолей молочных желез.
Оба примера указывают на связь экологии, социо-культурных традиций (питания) и собственно "биологической" или "внутренней" причины смертности человека, которые, таким образом, не могут рассматриваться как противостоящие полностью друг другу.
Новый подход к определению ВПЖ предложен в связи с обнаружением нового эффекта - "КОМПЕНСАЦИОННОГО ЭФФЕКТА СМЕРТНОСТИ". Он заключается в том, что при расчете интенсивности смертности для разных групп человека (или популяций различных видов животных) показатели "R" и "a" несколько варьируют, однако, для одного и того же вида все кривые проходят через ОДНУ ТОЧКУ на графике, что теоретически соответствует представлениям о наличии некоторой выделенной возрастной точки для вида. Для человека эта точка названа ВПЖ и соответствует 95 годам.
С учетом наличия некоторой выделяющейся общей точки в семействе кривых смертности для одного вида, закон Гомперца-Мейкема можно переписать теперь в виде: m(t) = A + M exp[a (t-В)],
где M и B - соответственно ордината и абсцисса такой выделенной точки.
Эти величины как раз и названы: В - видовой продолжительностью жизни, М - видовой интенсивностью смертности. Интересно, что обе величины оказались практически идентичными для мужчин и женщин, причем М соответствует примерно 50% гибели популяции данного возраста в течение года. Обращает внимание, что М в таком случае корреспондирует со "средними значениями" - например, с LD50 (летальная доза 50%) в токсикологии, определяющей величину действия фактора, вызывающего гибель 50% популяции. Можно было бы определить "В" как LT50 (летальный возраст 50%) - возраст, как своеобразный "летальный фактор", вызывающий гибель 50% популяции, однородной по данному фактору (то-есть, состоящей из равновозрастных особей).
Исследование компенсационного эффекта смертности и его осмысление выявляет ряд фундаментальных закономерностей. Так, наличие некоторой избранной точки в поздних возрастах говорит о наличии некоторых фундаментальных для вида механизмов, определяющих устойчивость организма (или механизмов старения), причем этот фактор должен быть связан с фундаментальными параметрами организма. Таким параметром некоторые считают, например, температуру, поддерживающуюся с высокой точностью в течение всей жизни. Соответственно, определяющими скорость старения механизмами считают, например, зависимые от температуры мутации (прежде всего точечные повреждения, вызываемые хаотическим температурным движением молекул и образовывающимися свободными радикалами).
Возможность самого компенсационного эффекта смертности связана с тем, что снижение интенсивности смертности ведет к увеличению темпов смертности (наблюдается "компенсация") и наоборот. Это, в частности, проявляется в том, что смерть человека, наступающая от одной болезни, при ее ликвидации "компенсируется" за счет другой. Такой почти мистический феномен можно объяснить только с достаточно фундаментальных позиций - смертность организма не складывается из смертностей от разных заболеваний, а распределяется по ним - то-есть, смертность рассматривается как вероятностный процесс снижения устойчивости сразу ко всем воздействиям, а не к отдельным из них. Это определяет, в частности, не эффективность чисто гериатрического подхода к решению проблемы увеличения ПЖ. Действительно, развитые страны столкнулись с феноменом все возрастающего значительного вложения средств в медицину пожилых при мало-заметном результате в отношении повышения ПЖ пожилых и старых лиц.
В связи с феноменом компенсационного эффекта смертности в геронтологию при обсуждении ПЖ вводятся представления системологии и кибернетики, как раз и описывающие общие характеристики систем, в частности, с позиций теории надежности и пр. При этом сама сложность системы и принципиально вероятностный механизм первичных отказов на любом уровне определяет нарастание со временем интенсивности отказов и выход всей системы в состояние "неспецифической уязвимости", что и ведет к "смерти от внутренних причин" или подставляет организм под удар любой внешней причины. Интересно, что оригинальные отечественные работы Гаркави и соавторов по адаптации как раз и предполагают возможность выхода организма в состояние неспецифически уязвимое или в состояние мало уязвимое, при этом в последнем случае наблюдается как "самопроизвольное" излечение от многих заболеваний, так и явные симптомы омоложения.
С другой стороны, разрабатываются тесты на диагностику такого неспецифического состояния уязвимости ко многим факторам внешней среды, включая травматизм, примером чего служит определение уровня сахара в крови, повышение которого как раз и указывает на выход организма в опасное состояние.
Таким образом, только весьма условно можно говорить о разделении факторов, влияющих на старение, на "внешние" и "внутренние", и признавать "биологическую основу старения" как чисто внутренний феномен организма.
Если говорить строго, то не удается определить ВПЖ как некоторый возраст, типичный для данного вида - для разных субпопуляций данного вида он оказывается разным. Для полевой мыши эта разница для весенних и осенних генераций составляет сотни процентов (различие в несколько раз и существование двух ВПЖ); для земноводных при различной температуре окружающей среды эта разница составляет уже тысячи процентов.
Кроме того, зависимость ПЖ от температуры для нетеплокровных довольно гладкая, что означает, что ВПЖ для них - не число, а функция то-есть ряд чисел, или ЗАКОН распределения.
В таком случае, правильнее говорить о ВПЖ как о законе распределения (только в ряде случаев вырождающемся в число) и определять параметры этого закона, а не как о некотором мифическом числе, которое не удается определить сколь-либо вразумительно ни с каких позиций.
Подход оценки МПЖ как характеристики особой точки пересечения нескольких графиков интенсивности смертности для одного вида на настоящий момент, видимо, является наиболее научным определением МПЖ, но весьма трудоемким и, видимо, мало пригодным для экспериментальных целей у животных.
ЕДИНАЯ СУЩНОСТНАЯ СИСТЕМНАЯ МОДЕЛЬ СТАРЕНИЯ
Таким образом, старение на современном этапе развития науки должно рассматриваться с достаточно общей методологической точки зрения, поскольку объективно принадлежит к феноменам наиболее общего типа – таким, как жизнь и смерть, бытие и сущность развития Природы в целом и т.п.
Современный системный анализ как методологический подход к старению, позволяет преодолеть изъяны прежних методов изучения старения как явления: метафизичность механистического рассмотрения как следствия какого-то конкретного физико-химического процесса или даже жесткой запрограммированности гибели организма, с одной стороны, и чрезмерной общности философских трактовок, не позволяющих сущностно наполнить и предметно анализировать этот процесс, что как раз и важно для практики. Смыкание фундаментальной и практической отраслей человеческой деятельности в настоящее время, реализованное в новой методологии - системеном анализе, позволило выработать достаточно полный, общий и конкретно наполненный метод изучения старения как глобального феномена жизни вообще и человеческой жизни в частности.
Системный подход к процессу старения включает формулирование глобального закона старения как общего феномена, что является определением старения и формулирует причину старения, которая принципиально должна быть определена на уровне достаточной абстрактной всеобщности: старение – глобальное снижение жизнеспособности (устойчивости ко всем формам повреждений), что ведет к повышению общей смертности целостного организма со временем в течение всей его жизни.
Общая причина старения в целом достаточно понятна на современном уровне обобщения – это дискретность существования организмов, ведущая к отграниченности их от внешней среды и приводящая к принципиальной недостаточности вследствие этого любых внутренних (следовательно, ограниченных пределами организма) механизмов самосохранения при принципиально неограниченном влиянии по качеству и силе со стороны внешних воздействий всего Мира. Это глобальное противоречие "целого и части" ведет к эволюции и развитию жизни, но включает изменение всех форм, а недостаточность изменчивости на индивидуальном уровне ведет к старению и смерти. Общность морфофункциональной организации особей конкретного вида придает им и общность сроков жизни, темпов старения и главных для каждого вида механизмов старения.
Системный анализ позволяет рассматривать старение с нескольких глобальных точек зрения, тем самым выявив 4 глобальных типа старения, как отражение принципиально однонаправленно действующих механизмов старения, что подробнее изложено ниже в отдельной главе:
а)"загрязнение" систем со временем как следствие принципиальной недостаточности открытости любых отграниченных от среды систем, даже если они самообновляются внутри себя;
б) принципиальная недостаточность сил отбора для сохранения только "нужных" структур в пределах данной системы, если информация для самосохранения имеется только внутри системы;
в) снижение количества любых необновляющихся структур в системе - а для большинства (сверх)сложных организмов всегда имеются необновляющиеся структуры на всех уровнях их организации;
г) регуляторные влияния в сложных системах, вызванные самыми различными причинами общего и частного характера, в том числе снижением давления эволюционного отбора с возрастом (после полового созревания) на направление и гармоничность систем регуляции; наличие конечных программ развития (роста, дифференцировки тканей, полового развития, иммунитета и т.п.).
Выделение 4-х типов старения позволяет ориентироваться в степени выраженности различных сторон процесса старения у каждого индивида и выбирать те или иные воздействия для геропрофилактики, биоактивации и продления жизни.
Системный подход позволяет количественно моделировать различные теории старения, причем нами выдвигается обязательное требование – сущностной наполненности модели, что позволяет сразу говорить о конкретных механизмах возможной реализации на биологическом уровне. Это, в частности, позволило нам подробно разработать новую иммунную модель старения, отражающую взаимоотношение пролиферирующих клеточных популяций различного типа и снижение потенциала клеточного роста с возрастом (что будет подробно рассмотрено в отдельной главе).
Важнейшим требованием системной теории старения является обязательный учет границ действия той или иной "теории" старения, на деле отражающей частные механизмы старения определенной группы клеток, органов, отдельные общие закономерности процесса старения и пр. – принцип иерархичности в рассмотрении механизмов и законов старения как глобального процесса.
Наконец, центральным положением системной теории старения является учет глобального единства целостного организма, целостности всех сторон его деятельности и изменений в течении жизни, а также принципиальное значение того факта, что, практически, феномен старения занимает важное место только в жизни человека и чисто человеческие качества (психические и социо-культурные) имеют едва ли не основное значение в рассмотрении старения вообще. Последнему обстоятельству благоприятствует то, что в природе в целом встречается удивительное разнообразие в сроках жизни и конкретных механизмах старения для каждого отдельного вида, что, однако, только подчеркивает то, что старение выраженно зависит от конкретного "устройства" организма и изучение старения во многом должно ориентироваться в первую очередь на человека. Именно человеческие особенности (культурные, религиозные, технические и пр.) и ограничивают возможные типы вмешательства в процесс старения, и открывают новые пути для него.
В целом, сам интерес к феномену старения для человека означает, что он вышел за пределы биологической природы и требует уже иных условий и форм существования. Фактически, речь идет о формировании новых путей эволюции человека, новых форм его жизнедеятельности и, возможно, нового типа физического тела, новых условий жизни и новых отношений со средой.
Таким образом, разрабатываемая нами системная сущностная теория старения является наиболее адекватным методом рассмотрения старения в настоящее время; она позволяет анализировать старение на всех уровнях: уровне общих закономерностей и понятий, уровне количественного моделирования, уровне сущностного наполнения биологическим материалом. При этом сохраняется общее видение проблемы и целостности организма со всеми его иерархическими уровнями организации. Эта теория принципиально открыта для развития и позволяет не отвергать уже имеющиеся теории старения, а органически вбирать их, рассматривая то место в целостной картине процесса старения, которое они объективно описывают.
Содержательная сторона отдельных достаточно разработанных частей этой общей системной сущностной теории старения и основанных на ней способов профилактики старения и подходов к биологической активации организма описываются в отдельных публикациях.
ЛИТЕРАТУРА
Гаврилов Л.А., Гаврилова Н.С.Биология продолжительности жизни.М:МОИП.Наука.1986.
Гвишиани Д.М. п/ред. Системные исследования. Ежегодник. ИСА РАН. М.:1996.
Донцов В.И. Иммунобиология постнатального развития. М.:МОИП. РАН. Наука.1990.
Донцов В.И. Применение теории гиперцикла для анализа процессов межклеточной регуляции пролиферации тканей: доказательства существования специализированной клеточной системы регуляции пролиферации тканей//Успехи современной биологии.1986.Т.101.Вып.1. С.18 -29.
Донцов В.И.,Крутько В.Н.,Подколзин А.А. Старение: механизмы и пути преодоления. М.: Биоинформсервис. 1997.
Комфорт А. Биология старения. М.:Мир.1967.
Крутько В.Н., Мамай А.В., Славин М.Б. Классификация, анализ и применение индикаторов биологического возраста для прогнозирования ожидаемой продолжительности жизни// Физиология человека. 1995. N 6. С.42.
Платон. Диалоги.М. П/ред. А.Ф.Лосева. М.:Мысль.1986.
Подколзин А.А.,Донцов В.И. Старение, долголетие и биоактивация. М.,1996.
Комментарии
ПРАКТИКА ПЕДИАТРА



